山西吕梁山草本群落对模拟增温的响应及与环境因子的关系
2020-08-04杨晓艳秦瑞敏张世雄徐满厚
杨晓艳,秦瑞敏,张世雄,温 静,鲁 钰,徐满厚
(太原师范学院,山西 晋中 030619)
【研究意义】气候变化通过改变植物光合作用、土壤酶活性等对陆地生态系统的结构和功能产生重要影响[1-2]。生态脆弱区、中高纬地区地理过渡带、山地高海拔地区作为全球气候变暖信号的放大器,其植物群落对气候变暖的响应更为敏感、迅速[3-4]。物种的分布是多个生态过程的产物,在不同尺度下,植被的群落组成及特征因生境差异均会出现一定的异质性,而山地的垂直变化将直接导致环境因子发生改变,进而影响山地植被的物种组成和群落结构。林下草本层群落因拥有较高的物种丰富度[5],是山地森林群落的重要组成部分。但目前对于山地森林群落的研究多集中于乔木及灌木部分,对林下草本层植物方面的研究并不充分[6]。因此,研究山地林下草本群落对模拟增温的响应极为重要。物种多样性和生物量是生态与环境科学领域研究的热点问题。物种多样性是促进经济发展、维持生态和谐的基础[7],也是衡量该地区生物资源丰富程度的标准;生物量是研究植物净初级生产力的基础,是衡量种间竞争的重要指标[8],也是生态系统结构与功能特征的重要表现。【前人研究进展】现今,多样性和生物量在全球变暖背景下的空间分布格局、对温度升高的响应及其作用机制成为生态学和地理学研究的重要议题[1],但对于该方面的研究多集中于单个山体或区域,但作为研究植物群落演替与稳定性的关键部分[9],多样性和生物量对深刻理解生态系统的结构和功能起到重要促进作用[10]。【本研究切入点】吕梁山位于黄土高原东麓,水土流失严重,是典型的生态环境脆弱区,其生态系统结构和功能对气候变化的响应极为敏感,然而目前有关吕梁山的研究多集中于山地植被群落的海拔梯度格局[11-12],运用系统论的观点及方法对吕梁山草本群落进行模拟增温方面的研究几乎未见报道。【拟解决的关键问题】因此,本文采用OTC野外模拟增温方法,在吕梁山林下草本层设置不同梯度增温,基于样地内草本植物高度、盖度、密度与频度等信息,结合所获取的环境因子,对其物种多样性及地上生物量随环境因子的变化进行比较与分析,以探究模拟增温对不同海拔梯度下山地草本群落发展情况的影响,为增温引起的植被退化提供有效保护与科学依据。
1 研究方法
1.1 研究区概况
吕梁山(35°41′~39°40′N、110°15′~112°20′E)位于山西省西部,地处黄土高原东麓(图1),呈南北走向,地跨忻州、吕梁和临汾3市,属大陆性半干旱季风气候,土壤类型垂直分布明显,从低海拔到高海拔依次分布有棕壤、褐土、草甸土及山地草甸土4个土类。此外,吕梁山脉纬度地带特征明显,可大致分为北段管涔山(38.25°~39.05°N、111.76°~112.9°E,海拔1310~2770 m)、中段关帝山(37.33°~38.33°N、110.3°~111.3°E,海拔1300~2830 m)和南段五鹿山(36.38°~36.63°N、111.03°~111.3°E,海拔1135~1950 m)。在管涔山实验区,土壤pH平均值为5.63,主要分布有寒温带针叶林,典型树种为云杉(Piceaasperata)和华北落叶松(Larixprincipis-rupprechtii);在关帝山实验区,土壤pH平均值为6.08,植被类型为温带针阔叶混交林,主要树种为白桦(Betulaplatyphylla)和云杉(Piceaasperata);在五鹿山实验区,土壤pH平均值为7.73,植被类型以暖温带阔叶林为主,典型树种为油松(Pinustabuliformis) 和辽东栎(Quercusliaotungensis)。

图1 吕梁山实验样地区位
1.2 样地设置
试验样地位于吕梁山系的五鹿山、关帝山和管涔山,对3座山均设置3个海拔梯度(五鹿山:1318、1459和1602 m;关帝山:1900、2001和2179 m;管涔山:2222、2395和2571 m),采用随机区组方法,选取物种分布均匀、人为干扰小的林下草本层植物进行样地布设,各样地面积为35~40 m2,共计9个样地。此外,对每个样地均设置对照(CK)、低度增温(OTC1)和高度增温(OTC2)3种处理,每种处理3个重复,样方间距1~3 m。需要指明的是,因关帝山低海拔地区(37°52′N、111°27′E,海拔1900 m)OTC2装置全部丢失,导致该样地高度增温处理3组数据缺失。
其中,增温样方采用无需电力供应的开顶式增温箱(Open-top chamber,OTC,图2)进行模拟增温处理,箱体为半封闭式正六边形,采用有机玻璃(透光率90 %以上)构建,厚度为4 mm,各斜边与地面夹角均为60°,OTC1和OTC2 2种增温装置的顶面边长均为25 cm,底面边长分别为50、75 cm,垂直高度分别为40、80 cm;对照样方(CK)不进行增温处理,植被处于自然生长状态,取样面积为1 m×1 m。

图2 2种增温装置
1.3 植被特征调查
于2018年8月(此时已进行2年的增温实验)从北向南依次对管涔山、关帝山和五鹿山各样地草本群落植被特征进行调查,测定指标包括各物种的名称、数量、频度、盖度和高度。实验采用0.5 m×0.5 m 网格样方框进行测量,并根据样方框大小自制网格尺寸为0.1 m ×0.1 m 的铁丝网,共计25个网格。此外,为避免样地破坏,在临近样地同种植被区域内随机选取长势相近的样方267个,采用网格法对各物种进行高度、盖度的测量,并将其刈割后运回实验室。各物种名由《中国植物志》得出。
1.4 环境因子调查
于2018年8月在距地面10 cm高度处架设带有防辐射罩(RS3-B)的空气温度传感器(S-THB-M008)进行空气温度的测定;在距地面10cm深度处,埋入便携式土壤温度传感器(S-TMB-M006)进行土壤温度测定,并采用数据采集器(H21-USB)以1 min为时间间隔进行数据存储。将0~10 cm深度的土样运回实验室自然状态下风干,并采用60目土筛将根系等杂质筛除,将土样与蒸馏水采用土水比1∶2.5进行配制,充分振荡并静置20 min后采用PH仪(PHSJ-4F型)与电导率仪(DDSJ-308型)分别进行土壤pH与电导率的测定。
1.5 数据处理
(1)α多样性指数包括 Simpson指数、 Shannon指数、 Pielou均匀度指数和 Patrick丰富度指数,计算公式如下:
(1)
R=S
(2)
(3)
(4)
(5)
(6)
式中:IV为重要值,rh为相对高度,ra为相对多度,rc为相对盖度,rf为相对频度;R为Patrick指数;H′ 为Simpson指数;H为Shannon指数;E为Pielou指数;i为样地内的植物物种i,Pi为第i个物种的相对重要值,S为样地内植物种类的总和。
(2)选择草本植物地上生物量的各项影响因子作为自变量(包括植物平均高度、总盖度、物种数),以地上生物量为因变量,建立回归方程(ABG=1.98×H+3.28×N+15.93×C-4.03,其中,H为植物平均高度;N为物种数;C为总盖度),根据此拟合方程进行不同增温梯度下地上生物量估测。
(3)运用SPSS19.0软件对物种多样性指数与生物量随温度变化情况进行曲线估计,并采用OriginPro 9.1软件绘制回归分析图。
(4)在SPSS19.0软件中,使用GLM(General Linear Model)模型,对吕梁山环境因子(空气温度与土壤温度、pH值、电导率)与植被群落特征(物种多样性和生物量)在各山地、海拔、增温处理下的异质性进行多因素方差分析,探究在不同海拔梯度下吕梁山林下草本群落与各环境因子关系的显著性及交互效应。
(5)运用Canoco for Windows 4.5软件对植被群落特征进行DCA(Detrended Correspondence Analysis)分析,确定采用RDA线性模型对吕梁山不同增温处理下各环境因子与植被的关系进行约束性排序分析。
2 结果与分析
2.1 环境因子对模拟增温的响应
在OTC1和OTC2处理下,五鹿山空气温度分别增加了0.56 %和4.57 %,关帝山空气温度分别增加了-0.12 %和3.03 %,管涔山空气温度分别增加了0.33 %和0.39 %(图3)。总体看来,在不同处理下吕梁山空气温度分别增加了0.27 %(OTC1)和3.33 %(OTC2),且在OTC2处理下,增幅随纬度升高极显著减小(P=0.006),空气日较差在不同处理下均随海拔升高而降低。

图3 不同增温处理下吕梁山不同海拔空气温度随时间的变化
在OTC1和OTC2处理下,五鹿山土壤温度分别下降了0.61 %和增加了1.91 %,关帝山土壤温度分别下降了1.89 %和增加了0.5 %,管涔山土壤温度分别下降了0.54 %和1.36 %(图4)。总体看来,在不同处理下吕梁山土壤温度分别减小了1.01 %(OTC1)和增加了0.95 %(OTC2),且在OTC2处理下,增幅随海拔升高显著下降(P=0.035),土壤日较差在不同处理下均随海拔升高而降低。
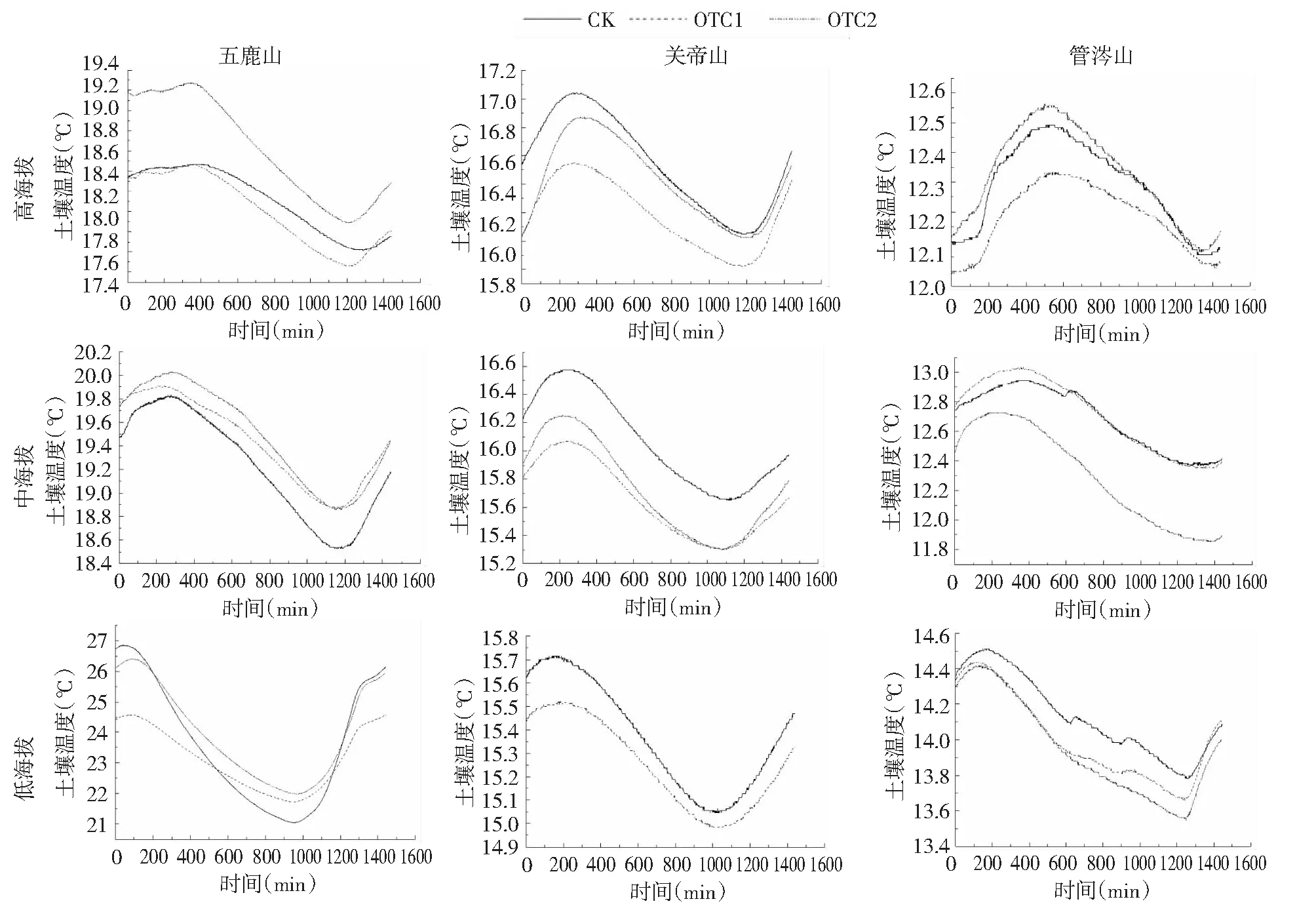
图4 不同增温处理下吕梁山不同海拔土壤温度随时间的变化
2.2 物种多样性对模拟增温的响应
Simpson指数、Shannon指数和Patrick指数随温度升高而下降(P>0.05),Pielou均匀度指数随温度升高略有上升(图5)。总体看来,Simpson指数、Shannon指数、 Pielou指数和Patrick指数在OTC1处理下分别增加了0.07 %、1.96 %、-8.62 %和0.00 %,在OTC2处理下分别减少了7.23 %、9.12 %、4.13 %和15.63 %。此外,物种多样性在不同山地、不同海拔梯度及山地与海拔共同作用下达到极显著水平,其中增温处理使Shannon指数和Patrick指数达到显著水平,海拔梯度与增温处理共同作用可使Simpson指数和Pielou指数达到显著水平,而不同山地、海拔、增温处理共同作用可使Simpson指数,Shannon指数和Pielou指数达到极显著水平(表1)。
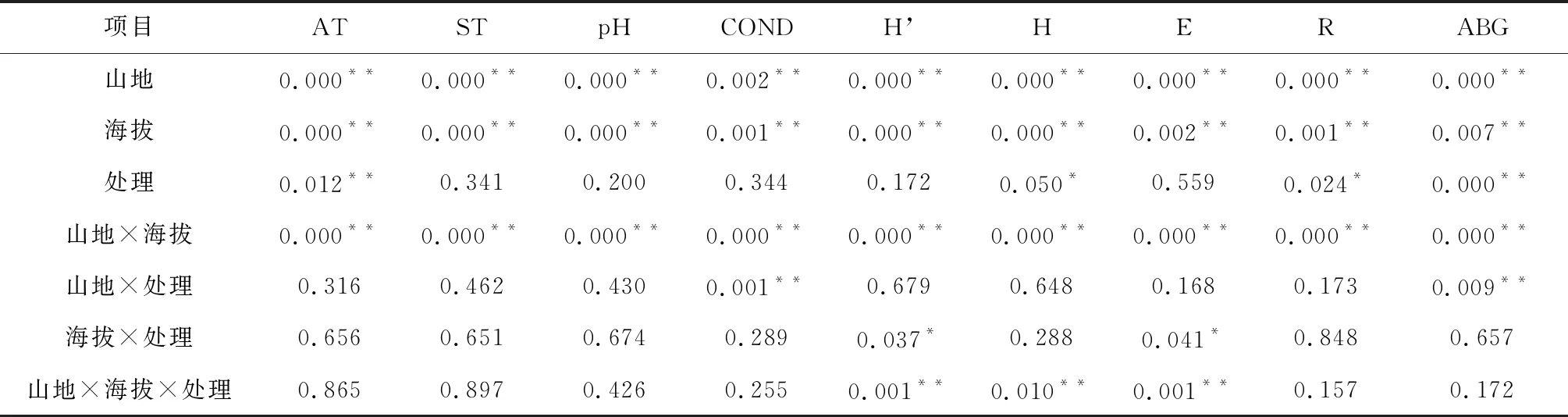
表1 吕梁山环境因子和植被在不同山地、海拔、增温处理下的多因素方差分析(P值)

图5 物种多样性指数随温度的变化趋势
在五鹿山,物种多样性指数随温度升高整体呈下降趋势(图6)。在低海拔,持续增温降低Simpson指数、Shannon指数和Patrick指数,而Pielou指数随温度升高略有增加;在中海拔,Simpson指数、Shannon指数和Patrick指数随温度升高先上升后下降,且Patrick指数在OTC1处理下达到显著水平;在高海拔,多样性指数随温度升高先下降后上升,其中OTC1处理可显著降低物种多样性,且Simpson指数、Shannon指数和Pielou指数为0 。
在关帝山,物种多样性指数随温度升高呈先上升后下降变化(图6),但整体呈下降趋势。在低海拔,OTC1处理可提高物种多样性(P>0.05);在中、高海拔多样性指数随温度升高呈先上升后下降变化,且Shannon指数和Patrick指数在不同处理间达到显著水平。
在管涔山,物种多样性指数随温度升高整体呈下降趋势(图6)。在低海拔持续增温使物种多样性降低;在中海拔,Simpson指数和Pielou指数随温度升高而下降,Simpson指数和Patrick指数随温度升高先上升后下降;在高海拔低度增温可增加植物物种多样性,过度增温使多样性降低(P>0.05)。
2.3 地上生物量对模拟增温的响应
吕梁山地上生物量随温度升高呈上升趋势(R2=0.2435,P<0.01,图7)。在五鹿山低海拔,增温可增加地上生物量,而中高海拔处,OTC1处理可增加地上生物量,OTC2处理抑制地上生物量增加;在关帝山各海拔梯度,增温均可增加地上生物量,但过度增温生物量增长减弱;在管涔山低海拔,增温对地上生物量无显著影响,在中海拔地区,OTC1处理可增加地上生物量,但OTC2处理导致地上生物量减少,且达到显著水平,在高海拔处生物量随温度升高呈增加趋势(图8)。此外,与物种多样性相比,地上生物量与环境因子的关系更为密切,在吕梁山不同山地、海拔、增温处理均可对草本植物地上生物量产生极显著影响(表1)。

图7 地上生物量随温度的变化趋势
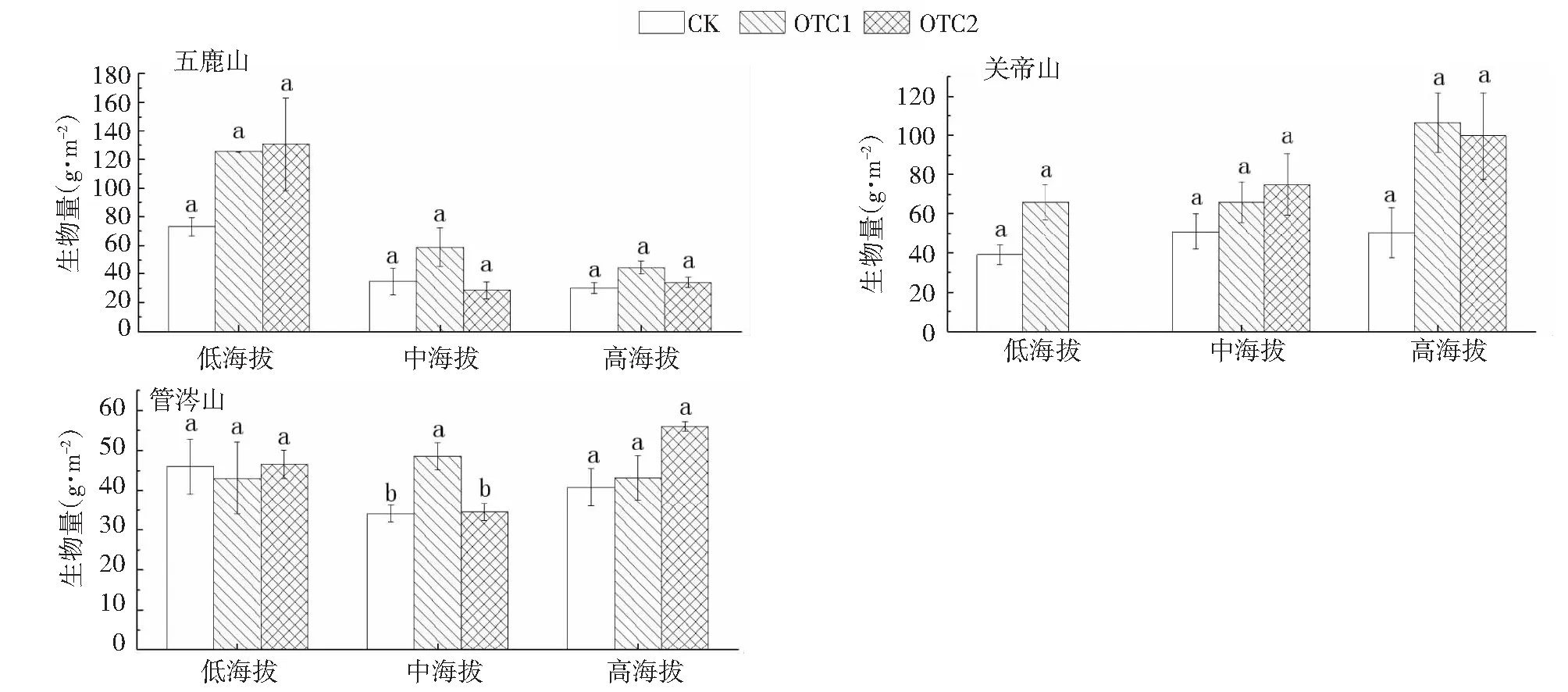
图8 地上生物量在不同处理下随海拔梯度的变化
总体而言,低度增温(OTC1)处理下,生物量增加了50.78 %,高度增温(OTC2)处理下,生物量增加了42.29 %,因此在吕梁山增温处理可极显著提高林下草本植物地上生物量。
2.4 不同增温处理下草本群落与环境因子关系
在不同增温处理下,草本植物物种多样性与各环境因子的相关性均未达到显著水平,但随温度升高两者相关性增强(图9)。物种多样指数与空气温度的关系在CK、OTC1处理下均呈负相关,而在OTC2处理下呈正相关;Simpson指数和Pielou指数与土壤温度的关系在CK、OTC1处理下呈负相关,在OTC2处理下呈正相关,Shannon指数和Patrick指数与土壤温度的关系在CK、OTC2处理下呈正相关,在OTC1处理下呈负相关;物种多样指数与土壤PH的关系在各处理下都呈负相关(除OTC2处理下的Pielou指数);物种多样指数与土壤电导率的关系在各处理下都呈正相关(除CK处理下的Patrick指数)。

图9 不同增温处理下吕梁山植被与环境因子的关系
在不同增温处理下,草本植物地上生物量与环境因子的关系整体呈正相关,且随温度升高两者相关性先增强后减弱(图9)。在CK条件下,地上生物量与空气温度、土壤温度及土壤PH均呈正相关关系,且与前两个环境因子(AT、ST)达到显著水平,而与土壤电导率呈负相关;在OTC1处理下,生物量与各环境因子间为正相关关系,且相关性均达到极显著水平;在OTC2处理下,生物量与各环境因子间也呈正相关关系,且极显著水平占75 %。
总体而言,草本群落物种多样性及生物量与环境因子的相关性随温度升高逐渐增强,RDA各排序轴的特征值总和在OTC1和OTC2处理下分别增加5.31 %和54.12 %(表2)。

表2 吕梁山不同增温处理下植被与环境因子关系的排序分析
3 讨 论
3.1 环境因子及其与植被的关系对温度升高的响应
由于开顶式增温箱(OTC)对空气的阻挡作用,导致室内风速降低、空气湍流减弱,在一定程度上改变了小室内下垫面情况。本研究表明,OTC模拟增温可提高室内空气温度,且增温箱高度与增温幅度呈正相关关系,这与众学者在各草地类型的研究结果相同[13-14]。但在土壤温度方面,本研究得出在低度增温处理下,土壤温度出现下降趋势,而高度增温使温度略为增加,经初步推断这可能与数据采集时的天气状况有关,在管涔山进行环境因子采集时遇雨水天气,降水量较大,而一些研究表明土壤温度与水分呈显著负相关关系[15],因此在低度增温情况下,土壤水分对土温的减弱程度超过了增温对其的增加程度,导致模拟增温使土壤温度出现下降的情况。
模拟增温通过影响土壤理化性质对植物的生长、发育等一系列生理过程产生影响,目前有关土壤理化性质对模拟增温响应方面的研究多集中于土壤有机质含量(C、N、P)、土壤酶活性与土壤微生物方面[16-17],关于模拟增温对土壤酸碱度影响方面的研究较少,且多集中于土壤pH随海拔梯度的变化格局方面[18-19]。本研究表明增温处理使土壤酸性增强,且土壤pH值与海拔呈极显著负相关关系。根据戴万宏等[20]对全国及6个地理区域地带性土壤表层有机质含量与pH的关系研究表明,两者呈现极显著负相关关系,且土壤pH变异可解释有机质含量总变异的18 %左右,因此,在模拟增温方面,初步推断增温处理抑制酶活性,有机质分解速度减缓,因而使土壤pH值较对照处理降低;在海拔梯度方面,由于随海拔升高山体土壤母质淋溶作用加强,土壤复合体吸附H+能力增强[18],从而使土壤pH值显著减小。
植物群落的空间分布格局是不同尺度条件下,各环境因子(如气候、土壤、地形等)综合作用的结果[21]。在区域及全球大尺度条件下,地带性气候条件(如温度、降水)被认为是决定植被类型空间分布的主导因素;而在景观及群落等小尺度条件下,非地带性环境因子(海拔梯度、土壤养分、坡度坡向等)将成为植被空间格局的主导作用[22-23]。本研究表明,随温度升高草本群落物种多样性及生物量与环境因子的相关性逐渐增强,且地上生物量在OTC1处理下与土壤pH呈极显著正相关关系。土壤酸碱度作为影响植物区系组成分异最主要的条件之一,其土壤pH值可综合反映土壤微生物活动、有机质的合成与分解及土壤养分保持的能力,进而对植物生物量生产及分配产生改变[24]。在本文由于温度升高加速有机质分解,从而提高植物地上生物量,而温度与土壤pH呈极显著正相关关系,因此吕梁山林下草本层群落地上生物量随土壤pH值增加极显著提高。
总之,模拟增温处理可提高空气温度,降低土壤温度与pH值,且在高度增温处理下,空气温度增幅具有负的纬度依赖性,土壤温度增幅具有负的海拔依赖性,并且在增温处理下可增强草本群落物种多样性及生物量与环境因子之间的关系。
3.2 物种多样性对温度升高的响应
增温可促进植物生长和发育[25],对植物群落的组成及演替产生影响。由于不同物种对增温的响应不同,全球变暖将打破原有种间竞争,使该群落组成发生变化[14,26-27],从而使物种趋于单一化发展[1]。在不同的草地类型,由于土壤条件和水热状况等不同,导致物种多样性对增温的响应各异,如在荒漠草原,增温可降低植物物种的多样性;在高寒草甸和矮嵩草草甸,短期增温可提高植被物种多样性;在东北羊草草甸,增温也可提高其多样性指数,且在增温第4年,植被物种多样性指数增加了16.8 %[28]。在群落演替方面,短期(1a)增温并不会改变群落种类组成,但增温效应会增强群落中建群种与主要伴生种的作用[29],长期过渡增温则会使植物物种趋于单一化发展[1],引起草地生态系统发生退化。笔者前期在吕梁山进行模拟增温实验的研究结果表明,在纬度较低的五鹿山,植物物种多样性在增温处理下呈递减趋势;在纬度较高的管涔山,适度增温可提高物种多样性,但过度增温多样性将受到抑制。本研究再次表明,位于黄土高原东麓的吕梁山短期内适度增温可提高物种多样性,但过度增温使多样性降低,这与众多学者在高寒地区的研究结果一致,因此可以看出,在较高海拔山地,低温同样是限制植物生长的关键因素之一,由于OTC的增温作用在一定程度满足了植物对热量的需求,使物种多样性增加,但过度增温超过植物所需最适温度,对植物生长起限制作用。
总之,短期内适度增温可提高植物物种多样性[30],但这种响应并不敏感[31],而长期模拟增温则降低植物群落物种数[32-33],引起草地生态系统发生退化。
3.3 地上生物量对温度升高的响应
温度升高通常会对植物生物量产生促进作用。相关研究表明,温度升高可使植物的生长期平均延长4.95 d[34-35],这将有助于草本植物获得更高的生物量。本人在吕梁山的研究也表明,林下草本植物地上生物量随温度升高显著增加。但由于生境及物种的不同,生物量对温度升高的响应各异,如在较干旱的温带地区,温度升高可显著提高地上部分的生物量;而在高寒地区,温度升高生物量将趋于地下部分[1,36]。
此外,增温对生物量的影响具有复杂性,增温可直接改变植物体酶的活性来影响其光合作用与呼吸作用[37],从而对植物有机质的累积产生影响,也可通过改变土壤氮素矿化速率和土壤水分含量[38],间接对生态系统净初级生产力产生影响。本研究得出,适度增温可显著提高地上生物量,过度增温生物量增长将减缓,这与众学者研究结果一致[1,13]。究其原因,主要是增温幅度较低时,可促进植物生长,使植物生物量得到累积;但当增温幅度过高时,将加剧OTC内土壤水分胁迫,使种间竞争加强,导致草本植物地上生物量增长减缓甚至降低。总之,增温会对植物生物量产生显著影响,短期内适度增温可促进植物生物量,但随着时间增长,生物量将减少[34]。
4 结 论
在环境因子方面,模拟增温对空气-土壤温度的影响差异较大。增温可提高空气温度,降低土壤pH值,此外,空气温度增幅具有纬度依赖性,土壤温度增幅具有海拔依赖性,且空气温度对模拟增温的响应更为敏感。在植物群落方面,适度增温可提高物种多样性与地上生物量,但当温度升高超过一定程度,这种促进作用会逐渐减弱甚至变为抑制作用,并且地上生物量对增温的响应更加敏感迅速。随温度升高草本植物与环境因子相关性增强,且过度增温可使物种趋于单一化发展,引起群落物种更替。
