北方草地及农牧交错区草地植被碳储量及其影响因素
2020-07-31辛晓平丁蕾程伟朱晓昱陈宝瑞刘钟龄何广礼青格勒杨桂霞唐华俊
辛晓平,丁蕾,程伟,朱晓昱,陈宝瑞,刘钟龄,何广礼,青格勒,杨桂霞,唐华俊
北方草地及农牧交错区草地植被碳储量及其影响因素
辛晓平1,丁蕾1,程伟1,朱晓昱1,陈宝瑞1,刘钟龄2,何广礼3,青格勒1,杨桂霞1,唐华俊1
(1中国农业科学院农业资源与农业区划研究所/呼伦贝尔草原生态系统国家野外科学观测研究站,北京 100081;2内蒙古大学生态与环境学院, 呼和浩特 010021;3锡林郭勒职业学院草原生态与畜牧业学院,内蒙古锡林浩特 026000)
【】草地生态系统在全球碳平衡中有重要的意义,草地植被碳库及其变化机制研究是植被生态学的重要命题。本文研究北方草地和农牧交错区草地植被碳密度及其空间格局,解析不同区域草地植被碳密度的关键影响因素,分析了气候、土壤、放牧等因素对地上地下植被碳库的相对贡献。基于2002—2009年北方草地及农牧交错带草地植被调查数据,结合同期MODIS/NDVI遥感影像和1﹕100万草地类型图,建立了我国主要草地类型的生物量估算模型;整合野外考察数据和前人研究结果,探讨了研究区地上地下生物碳库及其空间格局;基于研究区255个县级行政单元,分析了不同类型草地植被碳库与气候要素、土壤要素及家畜承载量的关系,应用一般线性模型(GLM)解析了不同影响因素对草地碳密度的相对贡献。(1)北方草地与农牧交错区草地地上平均生物碳密度为36.9 g C·m-2,地下生物碳密度为362.9 g C·m-2,地下生物碳密度高于地上10倍,均呈从东到西递减的趋势,频率分布图基本服从对数正态分布,不同草地类型的生物碳密度存在明显差异;(2)整个研究区及草原亚区、荒漠亚区、农牧交错亚区内,地上生物量与年降水量(MAP)呈极显著正相关、与年均气温(MAT)均呈极显著负相关,与土壤黏粒含量(Clay%)呈显著正相关、与土壤砂粒含量(Sand%)呈显著负相关,整个研究区家畜承载量与草地地上生物量之间呈极显著正相关;(3)一般线性模型(GLM)分析结果表明,年平均降水量(MAP)、年均气温(MAT)、土壤黏粒含量(Clay%)、放牧强度对地上生物量空间变异的解释率分别达到29.6%(<0.001)、5.8%(<0.001)、0.8%(<0.05)、1.3%(<0.001);地下生物量的空间变异主要来自于年降水量(MAP)、年均气温(MAT)、土壤砂粒含量(Sand%),对方差的解释率分别达到12.1%(<0.001)、6.8%(<0.001)、1.9%(<0.005),放牧强度没有明显贡献。气候条件尤其是年降水量是草地生物量碳库的主要影响因素,但对地上生物量影响更为明显;土壤质地对植被生物碳库也有显著贡献,尤其对地下生物量的影响更加显著;放牧强度只能解释地上生物量变化的1.3%、对地下生物量没有显著贡献,这一发现意味着气候对生物量碳库的贡献远大于放牧影响。
北方草地及农牧交错区;植被碳储量;气候因素;家畜承载量;土壤质地
0 引言
【研究意义】草地生态系统是重要的陆地碳库。由于草地生态系统所处环境的脆弱性,草地碳循环对全球变化的反应更加敏感,气候变化和不当的人类活动更易影响草地碳库的收支平衡,导致源汇逆转,因此,草地生态系统在全球碳平衡中占有重要位置[1-5]。【前人研究进展】自20世纪90年代后期,不少学者对我国区域到国家尺度的草地植被碳库进行了估算,但是不同研究给出的估算值存在很大差异。目前中国草地植被碳库(生物碳库)的估算值介于0.56—4.66 Pg C之间,其中地上生物量固定的碳占6%—17.2%。根据研究者、数据来源和计算方法,目前全国尺度上草地生物碳库估计值大致有4类:(1)NI等[6-8]估计的中国草地植被碳储量为3.05 Pg C(草地面积按照355万km2计),地上生物碳库储量0.134 Pg C;(2)FAN[9]利用78个地面实测样地、20世纪80年代全国草地资源普查数据,得到草地生物碳库储量为3.32 PgC(草地面积按照331万km2计),其中地上生物碳库储量0.393 Pg C;(3)Fang[10-11]、Piao[12]、Ma[13-14]等在不同时期利用20世纪80年代全国草地资源普查数据、2001—2005年全国近400个样地的实测数据、不同来源的遥感数据,估计中国草地生物碳库储量介于1.0—1.15 Pg C(草地面积按照331万km2计),其中地上部分占生物碳库储量的13.5—14.5%;(4)李克让等[15]估计的草地生物碳库储量为0.562 Pg C(草地面积按照167万km2计),是目前估计值中最小的。不同研究对区域草地生物量的估算也存在较大差异,其中Ni[16]、Fan[9]、Piao[12]、Ma[14]等对中国北方草地的估算分别为1.9、2.4、0.8、0.6 Pg C,最大值与最小值相差近4倍。作为受人类活动影响最剧烈的自然生态系统,草地植被碳库更多地受到全球变化的影响。在干旱到半湿润地区,降水是导致草地生态系统初级生产力动态变化的主要驱动因子[17-20],降水格局的变化(降水量的年际变化、季节分配及降水频率)会对植被生物量产生强烈影响[19,21-22]。温度也是影响植被碳库的关键要素,Valentini等[23]利用生态系统模型模拟结果表明,全球变暖会减小陆地碳汇的规模和数量,但LISKI等[24]指出,生态系统模型可能高估了土壤呼吸对温度的依赖性,全球变暖并不会促使陆地碳汇减少,相反,温度升高引发的负反馈会帮助碳在生态系统的累积。放牧利用是影响草地生态系统的碳循环关键人为因素之一。内蒙古锡林河流域羊草草原的研究结果表明,40年来过度放牧使表层土壤(0—20 cm)碳的贮存量降低了12.4%[25]。Ojima等[26]利用CENTURY模型研究大尺度草地生态系统碳循环时发现,放牧水平增加30%—50%导致未来50年温带草地将丧失大部分碳。【本研究切入点】位于北方草地及农牧交错区的草地生态系统是我国温带草地的主体,该区域自然条件严苛,草地生态系统物质循环和能量转换过程缓慢,对人类活动和气候变化反应敏感,是全球变化中一个很特殊的研究区域,草地植被碳库变化一直是该区域生态学研究的核心问题之一。草地植被生物碳库包括地上和地下两部分,草地生物碳库绝大部分在地下,在世界范围内,草地植被地下生物量与地上生物量的比值介于0.1—10.3。地上生物碳库主要取决于草地初级生产力固定的碳量,目前主要有2种主流估计方法:一是基于资源清查与遥感技术的统计推理,二是基于生态系统过程的模型模拟(如TEM、CENTURY等)。草地地下生物碳库的研究较地上部分困难得多,由于根系测量方法限制,大量的植物根系生物量数据难以获取,地下生物碳库一直是草地生态系统碳循环研究的薄弱环节。目前通常利用各类型草地地上部分与地下部分的比值(根冠比),估算草地地下植被的碳储量。【拟解决的关键问题】本文基于2002—2009年地面观测资料,结合同期遥感影像,估测北方草地及农牧交错区草地植被地上及地下部分碳密度和碳储量,分析草地植被碳密度的空间变异及其影响因素,为全面理解我国草地生态系统碳循环及其对全球变化的响应提供科学依据。
1 材料与方法
1.1 研究区域概况
研究区域包括了我国北方大部分草地及农牧交错带地区(34.85°—50.22°N,92.70°—127.8°E),在行政区划上包括内蒙古、黑龙江、吉林、辽宁、河北、山西、陕西、甘肃、宁夏、青海等10个省(区)的255个县级行政单元(旗、县和市辖区),自东至西由半湿润、半干旱向干旱区过度,相应分布着温性草甸、草甸草原、典型草原、荒漠草原及荒漠等地带性植被,以及荒漠区非地带性的高山植被。该区域是我国人口密度最高的草原地区(49人/km2),2001—2010年每年家畜存栏数在1.66—2.03亿羊单位之间波动。综合自然和经济特征,从东到西可以划分成农牧交错区、草原区、荒漠区等3个亚区域(图1)。
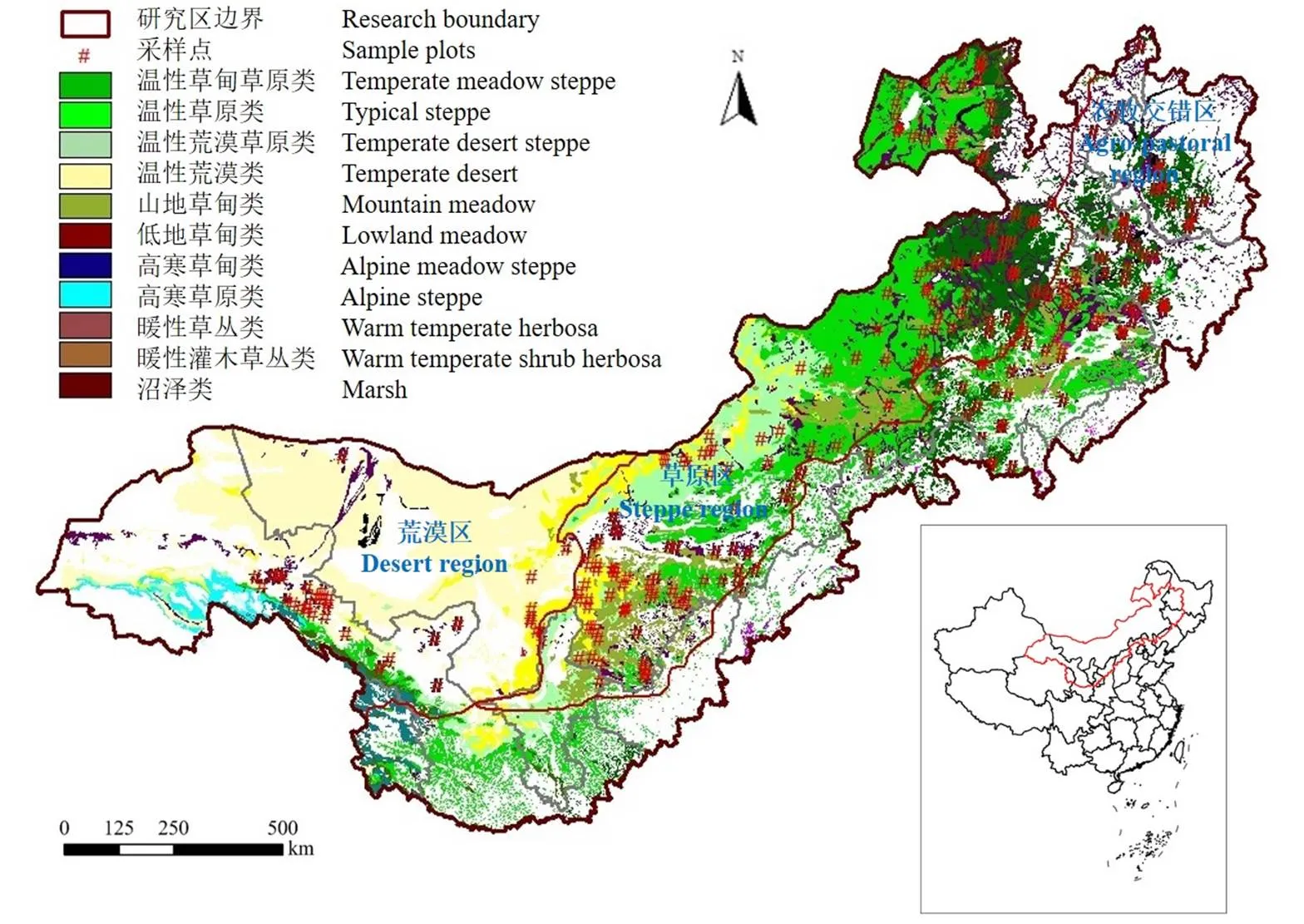
审图号:GS(2020)2229号
1.2 数据来源和预处理
本文所用遥感数据来自NASA免费提供的覆盖全国的1、2波段MOD09Q1产品。数据全部处理过程均采用国际上公认、可靠的方法,对MOD09Q1的8 d NDVI进行进一步的数据处理,包括图像配准、投影变换、填补像素、滤波去噪,并采用最大值合成法(MVC)提取研究区盛草期逐旬的NDVI最大值。NDVI植被指数数据集空间分辨率250 m×250 m,时间跨度2001年7月至2009年12月。本文涉及的遥感和空间数据处理分析均在ERDAS和ArcGIS软件下进行,地理投影采用双标准纬线等积圆锥投影(ALBERS),椭球体为KRASOVSKY,坐标系为Beijing 1954。
草地地面生物量数据来源于2002—2009年盛草期路线抽样调查(国家农业科学数据平台草地分中心支持)。草地生物量调查在北方草原最主要的、分布面积最大草地类型和群系选取比较均一(至少在周围2 km×2 km内相对均质)、对该草地类型的结构和生产力有足够的代表性的地段,布设1 km×1 km的样地,用GPS精确定位每个样地中心的地理位置,测定样地内物种组成、分布格局、地形和土壤条件。在样地内取5个样方,记录样方内分种多度、盖度和高度,用收割法测定地上生物量、土柱法测定地下生物量;根据样方所代表的草地类型在样地内面积比例,对5个样方的测量值加权平均,得到样地的平均地上生物量和平均草地群落盖度。高寒草甸和草原样方大小0.5 m×0.5 m,高寒荒漠、温性草甸和草原样方大小为1 m×1 m,温性荒漠样方大小为2 m×2 m或5 m×5 m。北方草地及农牧交错带共获得25个群系组469个样地、2345个样方的草地生物量数据(图1)。
气候数据来自中国气象局(http://cdc.cma.gov.cn/ home.do),包括2002—2009年全国712站的年降雨量、年均气温,应用反向距离加权法(IDW)插值形成1 km×1 km的气候栅格图像,通过空间分析统计得到研究区255个县在研究期限内的平均气温、平均降雨量数据。
本文应用家畜存栏数来表征放牧压力的影响。研究区各县2002—2009年家畜存栏数据来源于中国统计局农村社会经济调查队数据库。
1.3 草地地上生物量估算
模型方法是目前估算草地生物量及其动态的主要手段。本研究基于草地生物量地面调查数据和同期MODIS/NDVI遥感影像,结合我国1/100万草地类型图的分类系统,对我国主要草地类型建立了特异性草地生物量遥感模型,并在此基础上探讨中国草地生物量及碳储量时空格局。根据不同区域草地生物量与NDVI关系(图2-A),模型选择下列形式之一:
Y=a+b×NDVI,Y=a×ln(NDVI)+b或
Y=a×Exp (b× NDVI)

LM、MM、TMS、TS、TD、AM、AS分别代表低地草甸、山地草甸、温性草甸草原、典型草原、荒漠、高寒草甸、高寒草原
研究区469个有效样地中,313个用于建立模型,156个用于模型精度验证。结果表明草地生物量估测数据与实测数据有较好的符合(图2-B)。本文采用此估测数据开展植被碳储量研究。
1.4 统计分析
利用模型估测的草地地上生物量、不同类型草地的根冠比计算得到草地地下生物量。生物量采用国际上常用的转换率0.45统一以碳密度(g C·m-2)表示[10, 27]。各草地类型的平均地上、地下碳密度乘以类型面积得到碳储量。本文所涉及草地面积均来自1/100万草地类型图。采用ArcGIS软件空间分析模块,统计不同区域、不同草地类型及255个县域范围内平均草地碳密度。利用SPSS软件一般线性模型(GLM),分析气候因素和家畜数量对草地碳密度的影响程度。
2 结果
2.1 草地植被碳密度/碳储量格局
北方草地与农牧交错区草地地上平均生物碳密度为36.9 g C·m-2,地下生物碳密度将近地上生物碳密度的10倍(362.9 g C·m-2)。从分布格局看,地上、地下生物碳密度均呈从东到西递减的趋势,频率分布图基本服从对数正态分布(图3)。不同草地类型生物碳密度存在明显差异。温性草地地上生物碳密度从大到小依次是山地草甸、温性草甸草原、低地草甸、典型草原、荒漠草原及荒漠,分别是67.92、65.46、47.50、38.65、29.27、19.02 g C·m-2,不同类型间地下生物碳密度基本与地上生物量一致。高寒草地地上生物碳密度比温性草地偏低(高寒草甸38.43 g C·m-2,高寒草原17.31 g C·m-2),但由于高寒草地根冠比明显高于温性草地根冠比[28],其地下生物碳密度明显高于温性草地(高寒草甸1 456.2 g C·m-2,高寒草原482.71 g C·m-2)。但研究区域高寒草地只在荒漠区山地小面积分布,因此对区域碳储量贡献不大(表1)。
以研究区域面积115.28 km2计,北方草地与农牧交错区草地生物碳储量460.94 Tg,其中91.8%分布在地下碳库。对区域草地生物碳库贡献最大的是温性典型草原(29.7%),其余依次是温性草甸草原(19.9%)、低地草甸(18.2%)、温性荒漠(11.9%)、荒漠草原(8.4%)、高寒草甸(6.37%)。按照区域统计,草原亚区(51.94×104km2)草地生物碳储量215.00 Tg,农牧交错亚区(28.99 km2)及荒漠亚区(34.35 km2)分别为165.47 Tg、80.47 Tg,其中地上生物碳库分别占9.83%、9.01%、8.58%。
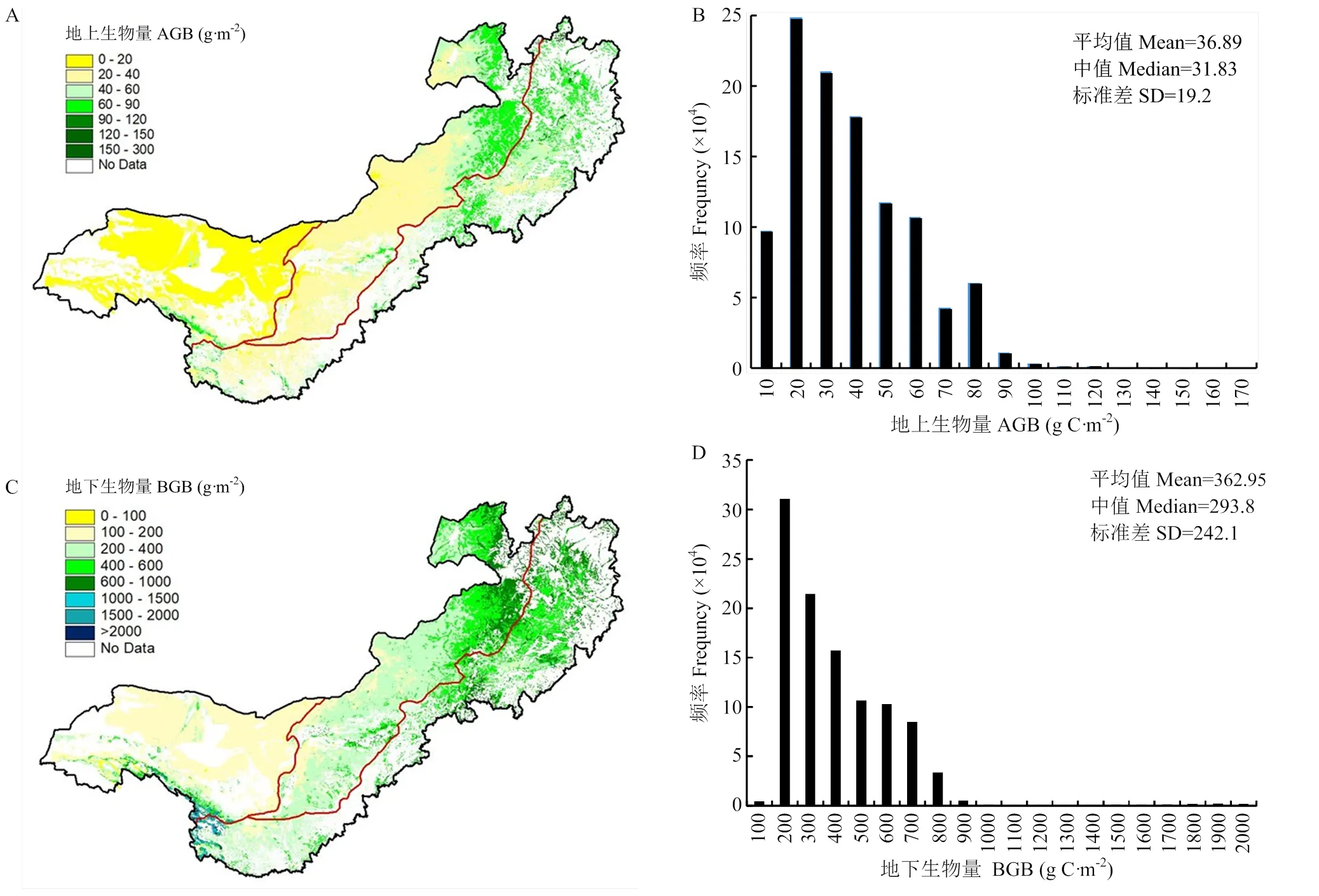
审图号:GS(2020)2229号
2.2 草地植被碳密度与气候、土壤及放牧压力的关系
对北方草地与农牧交错区及3个亚区内各县草地平均AGB与相应的气候、土壤及家畜存栏数进行双变量相关分析,结果如下:
(1)草原亚区、荒漠亚区、农牧交错亚区及整个研究区内,草地AGB与年降水量呈极显著正相关、与年均气温均呈极显著负相关(表2、图4)。在草原亚区与荒漠亚区,从东到西年均气温升高、年降水减少,地上生物量与年均气温、年降水的相关系数分别为0.25(<0.05)、0.64(<0.001)。农牧交错亚区从东北、内蒙到华北,随年均气温升高,气候从半湿润过渡到半干旱,草地生物量也随之降低。
(2)土壤质地与草地地上生物量之间存在显著相关性。草原亚区、荒漠亚区、农牧交错亚区及整个研究区土壤黏粒含量(Clay%)与草地AGB相关系数分别为0.54(2=0.2893,<0.001)、0.73(2=0.5373,<0.001)、0.46(2=0.2138,<0.001)、0.59(2=0.3479,<0.001),土壤砂粒含量与草地AGB相关系数分别为-0.65(2=0.4198,<0.001)、-0.63(2=0.3968,<0.01)、-0.23(2=0.0523,<0.01)、-0.42(2=0.1772,<0.001)。北方草地与农牧交错区以沙性土壤为主,研究区土壤黏粒含量只占5%—31.5%,而砂粒含量则高达31.5%—77.9%,在该区域,黏粒增加可以减少水分渗漏和散失,提高水分利用效率,从而具有较高的草地生物量。

表1 不同草地类型的植被碳密度和碳储量
TMS,TS,TDS,TD,LM,MM,AM,AS分别代表温性草甸草原、典型草原、荒漠草原、荒漠、低地草甸、山地草甸、高寒草甸、高寒草原
TMS, TS, TDS, TD, LM, MM, AM and AS represent respectively temperate meadow steppe, typical steppe, temperate desert steppe, temperate desert, lowland meadow, mountain meadow, alpine meadow and alpine steppe

表2 不同区域草地地上生物量(AGB)与环境因子的相关系数
***表示极显著相关(<0.001),** 表示强显著相关(<0.01),* 表示显著相关(<0.05)
**indicates extremely significant correlation (<0.01), * indicates strongly significant correlation (<0.01), *indicates significant correlation (<0.05)

A—D:地上生物量碳密度与平均年均气温的关系,E—H:地上生物量碳密度与平均年降水的关系,I—L:地上生物量碳密度与家畜承载量的关系,M—P:地上生物量碳密度与土壤质地的关系。DSE:羊单位
A-D: Relationship between above-ground biomass and mean annual temperature; E-H: Relationship between above-ground biomass and mean annual precipitation; I-L: Relationship between above-ground biomass and livestock capacity; M-P: Relationship between above-ground biomass and soil texture. DSE: Dry sheep equivalent
图4 不同区域草地地上生物碳密度(AGB)与气候、土壤及家畜存栏数关系
Fig. 4 Relationship between above-ground biomass (AGB) and climate, soil texture and livestock capacity
(3)家畜承载量与草地地上生物量之间的关系因区域而异,草原亚区、荒漠亚区、农牧交错亚区分别为不相关(Coef.=0.09,>0.05)、显著正相关(Coef.=0.45,<0.05)和极显著正相关(Coef.=0.34,<0.001),研究区总体呈极显著正相关(Coef.=0.25,<0.001)。草地生产力的大小决定区域家畜承载力,一般来说草地生产力越高的区域家畜承载量越大。草原亚区普遍超载,尤其是低生产力草原区域超载更严重,弱化了家畜承载量与草地生物量的相关关系。
3 讨论
3.1 草地植被碳库估计的不确定性
草地生物碳库绝大部分分布在地下[7, 29],由于根系测量方法限制,大量的地下碳循环数据难以获取,草地地下生物量的研究较地上部分困难得多,地下生物碳库是草地生态系统碳循环研究的薄弱环节[30-31]。方精云[32]研究和总结了近20年草地生态系统碳库变化,指出草地生态系统生物碳库、碳密度的估计存在很大的不确定性,不同研究者碳密度估算值相差可达5倍。除了所使用资料来源和估算方法的影响外,地下生物量难以准确获取也是不确定性的主要来源之一。
本研究使用2002—2007年野外采集的706对有效草地地下/地上生物量数据计算的不同类型草地根冠比[28],并与建国以来相关文献中不同草地类型根冠比进行了对比(表3)。可以看出,(1)本研究获得的不同草地类型根冠比整体上比YANG[33]、MA[14]的研究结果偏高,比FAN[9]的研究结果偏低。YANG、MA的根冠比考虑的是地下/地上活生物量比值,本研究考虑是全部地下生物量与地上生物量的比值。地下死生物量无法计入枯枝落叶生物碳库,土壤碳分析时这部分生物量也被剔除,未计入土壤碳库。如此计算,死亡根系存储的有机碳会被排除在地下生物碳库和土壤碳库之外,从而总体上低估草地生态系统地下碳库总量。尽管死亡根系已不具生命特征,我们仍然将这部分碳计入地下生物碳库。(2)温性草甸草原、典型草原、荒漠草原、低地草甸和山地草甸与其他研究者结果没有重大差异;高寒草甸、高寒草原根冠比与其他研究者相比显著偏高,但略低于FAN[9]通过大范围考察的计算结果;温性荒漠比大部分研究结果偏高,可能原因一是本研究中没有将温性草原化荒漠与温性荒漠做进一步区分,二是荒漠区以往研究多集中于灌木和小乔木荒漠,对本研究涉及的半灌木、小半灌木荒漠研究较少。
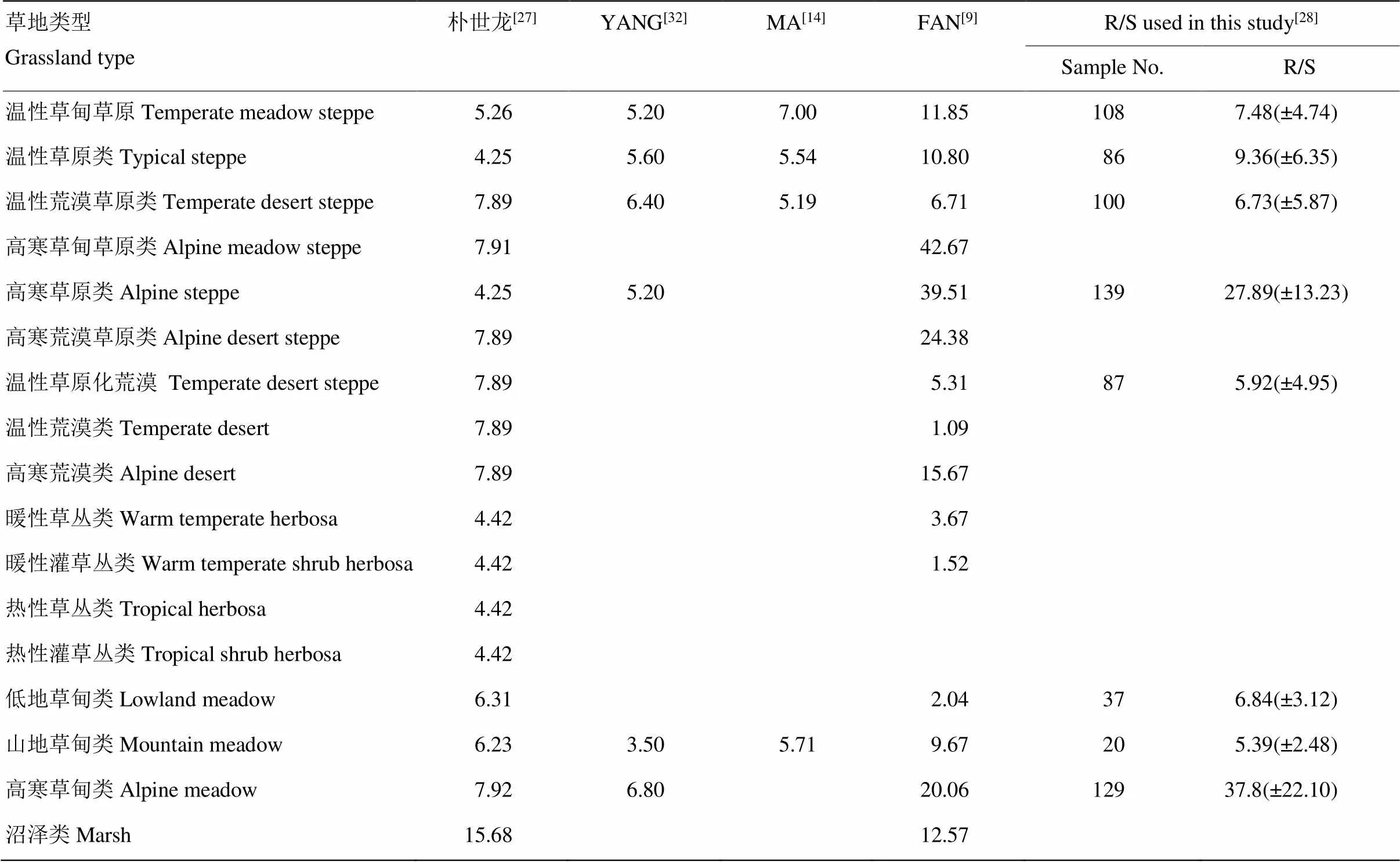
表3 不同研究中的草地类型根冠比
3.2 草地植被碳库的影响因素
草地是受人类活动影响最剧烈的自然生态系统,草地植被碳库不止受到气候、土壤等自然要素的影响,还与土地利用、植被管理等直接相关。我们对地上地下生物量及其影响因素的一般线性模型(GLM)分析(表4)结果表明,年降水量(MAP)、年均气温(MAT)、土壤黏粒含量(Clay%)、放牧强度对地上生物量空间变异的解释率分别达到29.6%(<0.001)、5.8%(<0.001)、0.8%(<0.05)、1.3%(<0.001)。地下生物量的空间变异主要来自于年降水量(MAP)、年均气温(MAT)、土壤砂粒含量(Sand%),对方差的解释率分别达到12.1%(<0.001)、6.8%(<0.001)、1.9%(<0.005),放牧强度的影响不显著。这表明气候条件尤其是年降水量是草地生物量碳库的主要影响因素,但对地上生物量影响更为明显;土壤质地对植被生物碳库也有显著贡献,但与气候要素相比,土壤质地对地下生物量的影响更加显著;放牧强度只能解释地上生物量变化的1.3%、对地下生物量没有显著贡献,这一发现意味着气候对生物量碳库的贡献远大于放牧影响,这一结果不同于以往将人类活动作为草地植被碳库关键驱动力的认识。
在干旱半干旱地区,降水是导致草地初级生产力变化的主要驱动因子,干旱年份和湿润年份的草地初级生产力可以相差±300%。放牧对草地生物量的影响比较复杂,适度放牧通过刺激植物的愈伤机制提高地上生物量生产、促进光合产物向地下分配,同时放牧家畜采食的干物质20%—60%以粪便形式返还到土壤中,加速有机碳周转速率;重度放牧会降低草地初级生产力,但对光合产物分配和根系周转的影响尚不明确。与草地地上植被一岁一枯荣相比,根系具有更长的寿命,孙元丰等[34]基于45篇相关文献整合分析表明,草原群落水平上根系周转在4—19年左右。因此,地下生物碳库具有更大的不确定性,其动态和机理也更为复杂,不止受气候条件影响,还与光合产物分配模式、根系的呼吸和死亡过程相关。但是目前,关于地下生态学的研究还比较有限,需要进一步的深入研究和探索。

表4 地上地下生物量与环境因子的一般线性模型分析结果
SS%变量所解释的方差比例Proportion of variances explained by the variable
4 结论
北方草地及农牧交错区是我国温带草地的主要分布区,其植被碳库、空间变异及影响因素是理解草原生态系统功能的核心问题之一。我们结合地面调查和遥感影响,估计了北方草地及农牧交错区草地植被地上及地下部分碳密度和碳储量,分析了草地植被碳密度的空间变异及其影响因素,主要结论认为:
(1)北方草地与农牧交错区草地地上平均生物碳密度36.9 g C·m-2,地下生物碳密度将近地上生物碳密度的10倍(362.9 g C·m-2),不同草地类型生物碳密度存在明显差异;
(2)地上生物量与年降水量呈极显著正相关、与年均气温均呈极显著负相关,整个研究区家畜承载量与草地地上生物量之间呈极显著正相关;
(3)气候条件尤其是年降水量是草地生物量碳库空间变化的主要影响因素,气候对草原地上生物量碳库变化的贡献率达到35.4%,对地下生物量碳库的解释率达到18.7%;土壤质地对植被地下生物碳库有显著影响,放牧强度对草原地上生物量碳库空间变化有显著影响,但是土壤质地和放牧强度对草原植被碳库空间变异的贡献率都低于2%。气候对生物量碳库的贡献远大于放牧影响。
[1] BOLIN B. Changes of land biota and their importance for the carbon cycle., 1977, 196(4290): 613-615.
[2] AJTAY G. Terrestrial primary production and phytomass., 1979: 129-181.
[3] ROY J, SAUGIER B, MOONEY H.. San Diego: Academic Press, 2001.
[4] BAAH-ACHEAMFOUR M, CHANG S X, CARLYLE C N, BORK E W. Carbon pool size and stability are affected by trees and grassland cover types within agroforestry systems of western Canada., 2015, 213: 105-113.
[5] ZHOU G Y, QIN L, JIE C Y, MIAO H, YAN Z L, DOUGLAS F, HUI H Y, LING F Y, CHENG Z B, HUI Z X. Effects of livestock grazing on grassland carbon storage and release override impacts associated with global climate change., 2019, 25(3): 1119-1132.
[6] NI J. Carbon storage in terrestrial ecosystems of China: estimates at different spatial resolutions and their responses to climate change., 2001, 49(3): 339-358.
[7] NI J. Carbon storage in grasslands of China., 2002, 50(2): 205-218.
[8] NI J. Forage yield-based carbon storage in grasslands of China., 2004, 67(2/3): 237-246.
[9] FAN J W, ZHONG H P, HARRIS W, YU G R, WANG S Q, HU Z M, YUE Y Z. Carbon storage in the grasslands of China based on field measurements of above- and below-ground biomass., 2008, 86(3): 375-396.
[10] FANG J Y, GUO Z D, PIAO S L, CHEN A P. Terrestrial vegetation carbon sinks in China, 1981-2000., 2007, 50(9): 1341-1350.
[11] FANG J Y, LIU G H, XU S L. The carbon cycle of terrestrial ecosystems in China and its global significance in china//. Beijing: Chinese Environmental Science Publishing House, 1996: 129-139.
[12] PIAO S L, FANG J Y, ZHOU L M, TAN K, TAO S. Changes in biomass carbon stocks in China's grasslands between 1982 and 1999., 2007, 21(2): 1-10.
[13] MA W H, FANG J Y, YANG Y H, MOHAMMAT A. Biomass carbon stocks and their changes in northern China’s grasslands during 1982–2006., 2010, 53(7): 841-850.
[14] MA W H, LIU Z L, WANG Z H, WANG W, LIANG C Z, TANG Y H, HE J S, FANG J Y. Climate change alters interannual variation of grassland aboveground productivity: evidence from a 22-year measurement series in the Inner Mongolian grassland., 2010, 123(4): 509-517.
[15] 李克让, 王绍强, 曹明奎. 中国植被和土壤碳贮量. 中国科学: 地球科学, 2003, 33(1): 72-80.
LI K R, WANG S Q, CAO M K. Carbon storage of vegetation and soil in China., 2003, 33(1): 72-80. (in Chinese)
[16] NI J, SYKES M T, PRENTICE I C, CRAMER W. Modelling the vegetation of China using the process-based equilibrium terrestrial biosphere model BIOME3., 2000, 9(6): 463-479.
[17] 王玉辉, 周广胜. 内蒙古羊草草原植物群落地上初级生产力时间动态对降水变化的响应. 生态学报, 2004, 24(6): 1140-1145.
WANG Y H, ZHOU G S. Response of temporal dynamics of aboveground net primary productivity ofcommunity to precipitation fluctuation in Inner Mongolia., 2004, 24(6): 1140-1145
[18] BAI Y F, HAN X G, WU J G, CHEN Z Z, LI L H. Ecosystem stability and compensatory effects in the Inner Mongolia grassland., 2004, 431(7005): 181-184.
[19] BAI Y F, WU J G, XING Q, PAN Q M, HUANG J H, YANG D L, HAN X G. Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau., 2008, 89(8): 2140-2153.
[20] 何楷迪, 孙建, 陈秋计. 气候要素和土壤质地对青藏高原草地净初级生产力和降水利用率的影响. 草业科学, 2019, 36(4): 140-152.
He K D, SUN J, CHEN Q J. Response of climate and soil texture to net primary productivity and precipitation-use efficiency in the Tibetan Plateau., 2019, 36(4): 140-152.
[21] 郭群. 草原生态系统生产力对降水格局响应的研究进展.应用生态学报, 2019, 30(7):2201-2210.
GUO Q. Responses of grassland ecosystem productivity to altered precipitation regime: A review., 2019, 30(7): 2201-2210. (in Chinese)]
[22] WALTER J, GRANT K, BEIERKUHNLEIN C, KREYLING J, JENTSCH A. Increased rainfall variability reduces biomass and forage quality of temperate grassland largely independent of mowing frequency., 2012, 148: 1-10.
[23] VALENTINI R, MATTEUCCI G, DOLMAN A, SCHULZE E-D, REBMANN C, MOORS E, GRANIER A, GROSS P, JENSEN N, PILEGAARD K. Respiration as the main determinant of carbon balance in European forests., 2000, 404(6780): 861-865.
[24] LISKI J, ILVESNIEMI H, MäKELä A, WESTMAN C J. CO2emissions from soil in response to climatic warming are overestimated -The decomposition of old soil organic matter is tolerant of temperature., 1999, 28(2): 171-174.
[25] LI L, CHEN Z, WANG Q, LIU X, LI Y. Changes in soil carbon storage due to over-grazing insteppe in the Xilin River Basin of Inner Mongolia., 1997, 9(4): 104-108.
[26] OJIMA D S, DIRKS B O M, GLENN E P, OWENSBY C E, SCURLOCK J O. Assessment of C budget for grasslands and drylands of the world., 1993, 70(1): 95-109.
[27] 朴世龙, 方精云, 贺金生, 肖玉. 中国草地植被生物量及其空间分布格局. 植物生态学报, 2004, 28(4): 491-498.
PIAO S L, FANG J Y, HE J S, X Y. Spatial distribution of grassland biomass in China., 2004, 28(4): 491-498. (in Chinese)
[28] 王道龙, 辛晓平. 北方草地及农牧交错区生态--生产功能分析与划分. 北京: 中国农业科学技术出版社, 2011.
WANG D L, XIN X P.-. Beijing: China Agricultural Science and Technology Press, 2011. (in Chinese)
[29] POST W M, EMANUEL W R, ZINKE P J, STANGENBERGER A G. Soil carbon pools and world life zones., 1982, 298(5870): 156-159.
[30] 贺金生, 王政权, 方精云. 全球变化下的地下生态学: 问题与展望. 科学通报, 2004, 49(13):1226-1233.
He J S, WANG Z Q, FANG J Y.Underground ecology under global change: problems and prospects., 2004, 49(13):1226-1233. (in Chinese)
[31] 吴伊波, 崔骁勇. 草地植物根系碳储量和碳流转对CO2浓度升高的响应. 生态学报, 2009, 29(1): 378-388.
WU Y B, CUI X Y. Responses of root carbon reserves and root turnover to experimental CO2enrichment in grasslands., 2009, 29(1): 378-388. (in Chinese)
[32] 方精云, 杨元合, 马文红, 安尼瓦尔·买买提, 沈海花. 中国草地生态系统碳库及其变化. 中国科学: 生命科学, 2010, 40(7): 566-576.
FANG J Y, YANG Y H, MA W H, ANIWAER M M T, SHEN H H. Ecosystem carbon stocks and their changes in China’s grasslands., 2010, 40(7): 566-576.
[33] YANG Y H, FANG J Y, MA W H, GUO D L, MOHAMMAT A. Large-scale pattern of biomass partitioning across China’s grasslands., 2010, 19(2): 268-277.
[34] 孙元丰, 万宏伟, 赵玉金, 陈世苹, 白永飞. 中国草地生态系统根系周转的空间格局和驱动因子. 植物生态学报, 2018, 42(3): 337-348.
SUN Y F, WAN H W, ZHAO Y J, CHEN S P, BAI Y F. Spatial patterns and drivers of root turnover in grassland ecosystems in China., 2018, 42(3): 337-348. (in Chinese)
Biomass Carbon Storage and Its Effect Factors in Steppe and Agro-pastoral Ecotones in Northern China
XIN XiaoPing1, DING Lei1, CHENG Wei1, ZHU XiaoYu1, CHEN Baorui1, LIU ZhongLing2,HE GuangLi3, QING GeLe1,YANG GuiXia1, TANG HuaJun1
(1Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences/National Hulunber Grassland Ecosystem Observation and Research Station, Beijing 100081;2Department of Environmental Sciences,Inner Mongolia University, Huhhot 010021;3Department of Grassland Ecology and Animal Husbandry,Xilingol Vocational College, Xilinhot 026000, Inner Mongolia)
【】The grassland ecosystem plays an important role in the global carbon balance. The study of grassland carbon pool and its driving force is a hot point of vegetation ecology. This study investigated the vegetation carbon density and its spatial pattern in the steppe and agro-pastoral ecotones of northern China. The major factors driving the spatial variation of grassland vegetation carbon density were identified, as well as the relative contribution of climate, soil texture, grazing intensity and other factors to the grassland vegetation carbon pool. 【】Using the survey data of the grassland vegetation in northern grassland during 2002 and 2009, combined with the MODIS/NDVI remote sensing data and 1:1 million grassland type map, the estimation model of above- and below-ground biomass in the main grassland types of northern China was established. Based on 255 county-level administrative units in the study area, the relationship between grassland vegetation carbon density and climate factors, soil texture and livestock carrying capacity were explored, and derived the relative contribution of different driving factors to grassland carbon density using the general linear model (GLM). 【】 (1) The average above-ground biomass (AGB) of the steppe and agro-pastoral ecotones of northern China was 36.9 g C·m-2, and the below-ground biomass (BGB) was 362.9 g C·m-2, nearly 10 times the AGB. Both the above- and below-ground biomass decreased from east to west, and followed logarithmic normal distribution. The biomass carbon density of grassland types was significantly different. (2) In the whole study region and steppe sub-region, desert sub-region, agro-pastoral sub-region, the AGB showed a significantly positive correlation with mean annual precipitation (MAP) and soil clay content (Clay%), a significantly negative relationship with the mean annual temperature (MAT) and soil sand content (Sand%). The AGB increased with livestock carrying capacity except in the steppe sub-region where were very heavily grazed. (3) General Linear Model (GLM) analysis indicated that the MAP, MAT, Clay% and grazing intensity explained 29.6% (<0.001), 5.8% (<0.001), 0.8% (<0.05) and 1.3% (<0.001) of AGB variation, respectively, and the MAP, MAT and Sand% contributed to 12.1% (<0.001), 6.8% (<0.001) and 1.9% (<0.005) to BGB variation, respectively, and the grazing intensity had minor contribution to BGB. 【】Climate factors especially MAP was the dominate driving factor of grassland vegetation carbon density, and its impact on AGB was more obvious than on BGB. Soil texture also had a significant contribution to the grassland vegetation carbon density, especially on the BGB. Grazing intensity explained only 1.3% of the AGB and had no impact on BGB. This finding indicated that the climate factors were major contributor grassland vegetation carbon density comparing with grazing intensity.
steppe and agro-pastoral ecotones in northern China; biomass carbon storage; climate; livestock capacity; soil texture

10.3864/j.issn.0578-1752.2020.13.022
2019-09-23;
2020-02-19
国家重点研发计划(2016YFC0500600,2017YFE0104500)、国家自然科学基金(41771205)、现代农业产业技术体系建设专项资金(CARS-34)
辛晓平,Tel:010-82109615;E-mail:xinxiaoping@caas.cn。通信作者唐华俊,E-mail:tanghuajun@caas.cn
(责任编辑 林鉴非,杨鑫浩)
