梁河家鼠鼠疫疫源地鼠疫噬菌体的分离及其流行病学意义
2020-07-28张海鹏钟佑宏段存娟吴鹤松
张海鹏,钟佑宏,段存娟,郭 牧,段 彪,李 伟,王 鹏,吴鹤松
梁河县位于云南省西部,地处东经98°09′-98°39′,北纬24°35′-24°56′,年平均气温18.28 ℃,年均降水量1 334 mm,是云南省家鼠鼠疫疫源地。自1870年有鼠疫的流行记载以来,1944年到1955年发生人间病例2 047例[1],之后进入微弱流行期,只在鼠间偶有分离到鼠疫菌。80年代鼠疫复燃,1990年于遮岛镇、九保乡再次暴发鼠间鼠疫,流行至2001年,并出现1例人间腺鼠疫[2],之后渐渐转弱,2007年在鼠间偶有流行。2008年至今,持续监测未发现鼠疫流行的相关信息,鼠疫流行进入静息期。噬菌体是一种具有严格宿主特异性只感染一种细菌或只寄居在宿主菌体内而不感染哺乳动物细胞的病毒[3-4]。梁河县家鼠鼠疫疫源地中有无鼠疫噬菌体的存在,鼠疫噬菌体在不同的宿主动物中携带率是否有差异,其对鼠疫的流行与静息是否有特殊意义。为此本研究对梁河县家鼠鼠疫疫源地曾流行过鼠疫的乡镇进行鼠疫噬菌体分离鉴定。
1 材料与方法
1.1标本来源 2017年选取梁河县4个曾流行过鼠疫的乡镇(九保乡、芒东镇、河西乡、遮岛镇)利用鼠铗法捕获的鼠类动物经鉴定后在无菌条件下解剖,取肠置于盛有10 mL改良PBS增菌液的培养管中,4 ℃冰箱保存待检。
1.2主要仪器设备、培养基和试剂 日立HT770透射电镜、0.22 μm除菌滤器、恒温气浴振荡培养箱;改良PBS增菌液、LB培养基(固、液)均按常规方法制备和保存。
1.3鼠疫噬菌体的分离[5-6]将增菌液用0.22 μm除菌滤器过滤,获得噬菌体原液,吸取10 mL加入到含有EV76菌悬液100 μL的5 mL LB液体培养基中,充分混匀,置培养箱28 ℃、220 r/min振荡培养18 h。取200 μL再次过滤的上清,加入100 μL EV76菌悬液充分混匀,倾倒双层平板,置28 ℃培养24 h后观察有无噬菌斑。
1.4鼠疫噬菌体的纯化 挑取单个噬菌斑加入到含有对数生长期鼠疫疫苗株EV76菌悬液300 μL的30 mL LB液体培养基中,重复进行单斑增殖6次,使所分噬菌体得到纯化。
1.5鼠疫噬菌体电镜扫描样本制备[5-6]制作EV菌双层平板,在表面滴加噬菌体,20 h左右形成大块噬斑,用接种环取下上层半固体,浸泡于200 μL生理盐水中1 h左右得到样本,经醋酸铀染色,80 kV,放大倍数1 000~200 000条件下电镜观察。
2 结 果
2.1不同乡镇分离鼠疫噬菌体结果 2017年梁河4乡镇共捕鼠338只,其中芒东镇捕鼠207只、九保乡捕鼠51只、河西乡捕鼠49只、遮岛镇捕鼠31只。共分离到29株鼠疫噬菌体,总分离率为8.58%。其中遮岛镇分离到5株鼠疫噬菌体,分离率16.13%,分离率最高,将近总分离率的2倍,河西乡分离到6株鼠疫噬菌体,分离率12.24%,九保乡分离到6株鼠疫噬菌体,分离率11.76%,芒东镇分离到12株鼠疫噬菌体,分离率5.8%为最低,详见表1与图1。
2.2不同鼠种鼠疫噬菌体的分离结果 本次调查捕获的338只宿主动物包括野鼠、家鼠及其它野生小兽共14种,其中黄胸鼠为优势宿主动物,共捕获234只,占此次捕获动物总数的69.23%,其次为臭鼩鼱,捕获率为12.82%。从234只黄胸鼠中分离到19株鼠疫噬菌体,分离率为8.12%;从43只臭鼩鼱中分离到8株鼠疫噬菌体,分离率为18.6%,详见表2与图1。
2.3所分噬菌体的噬菌斑 29株鼠疫噬菌体噬斑表现出多态性。有噬斑较大(直径≥2.0 mm)且透明(如143号)者,有噬斑中等(2.0 mm≥直径≥1.0 mm)且透明者(如337号),有噬斑较小(直径<1.0 mm)且半透明者(如72号与332号),详见图2。
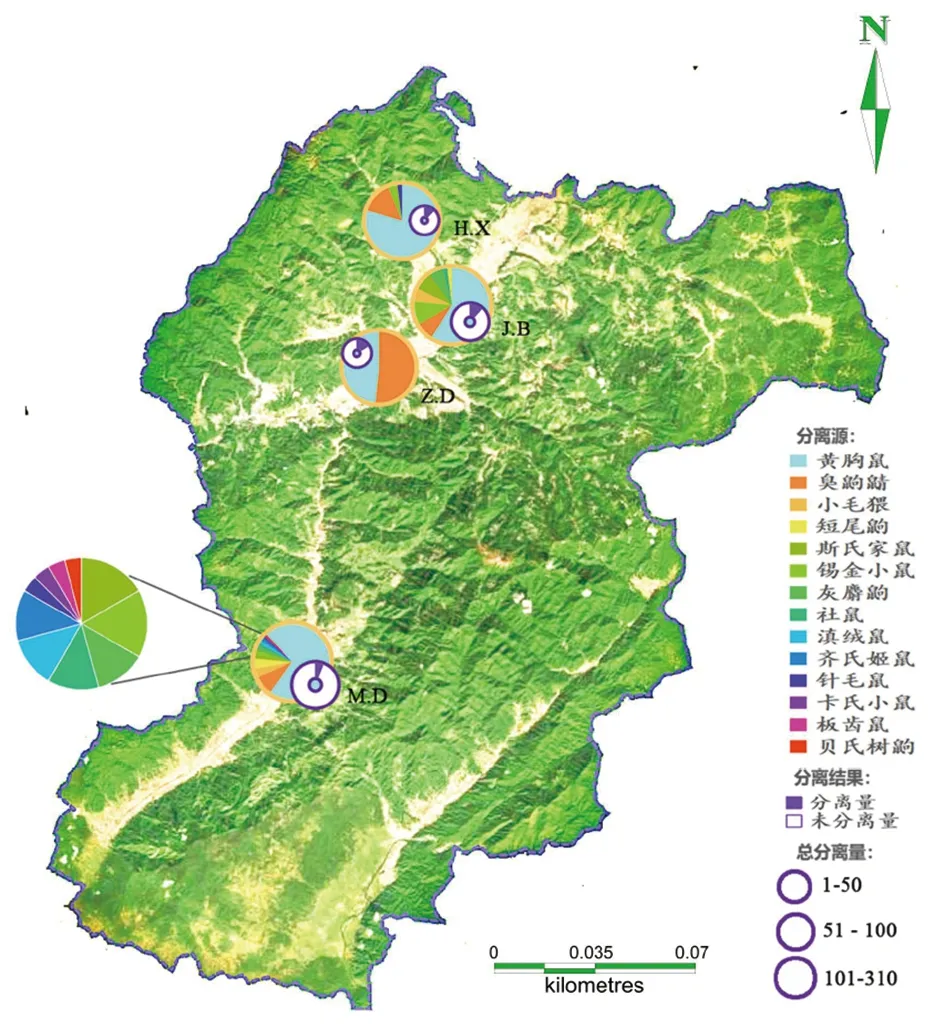
图1 梁河县4乡镇不同鼠种中分离鼠疫噬菌体地理图Fig.1 Geographic map of Yersinia pestis phage isolated from different rats in 4 towns of Lianghe County
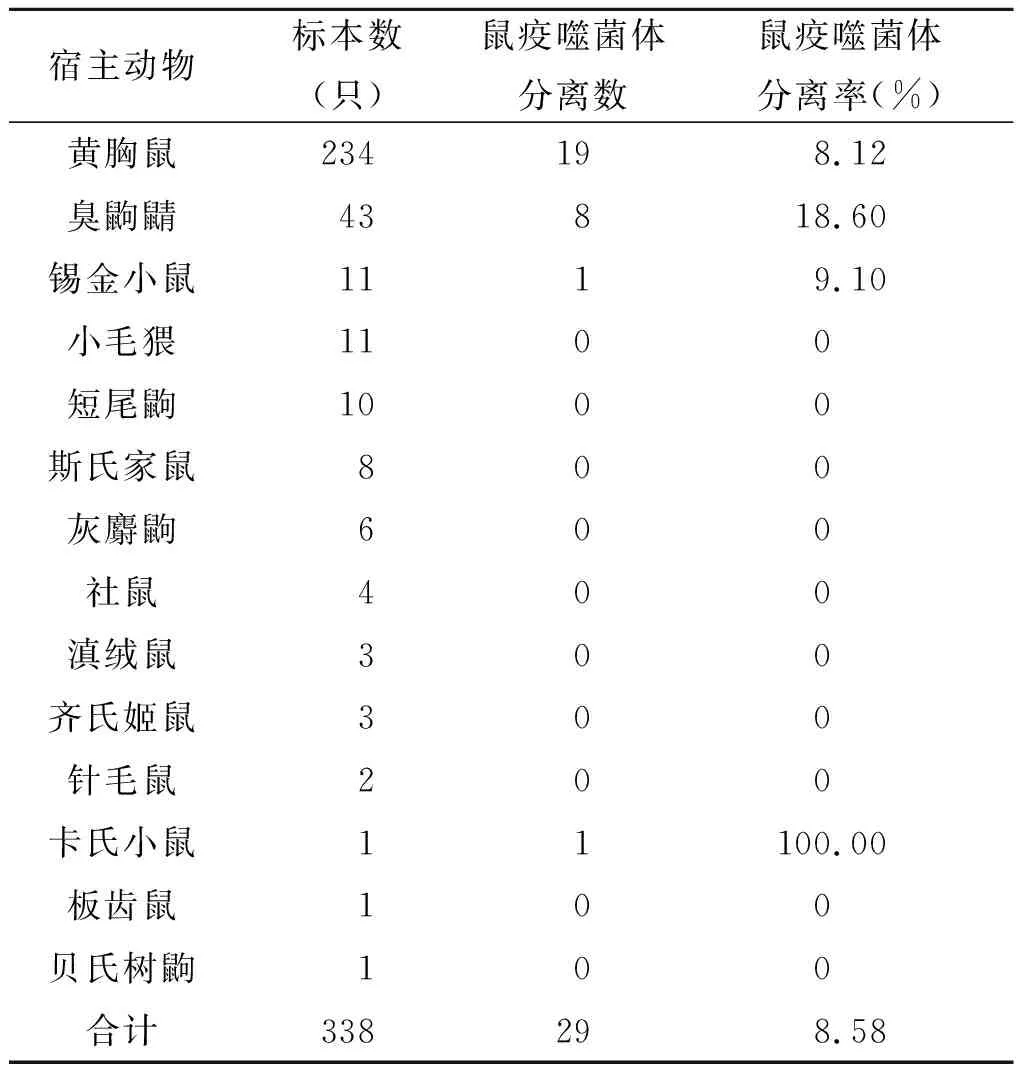
表2 梁河县不同宿主动物鼠疫噬菌体的分离结果Tab.2 Results of Yersinia pestis phage isolation in different host animals in LiangHe County
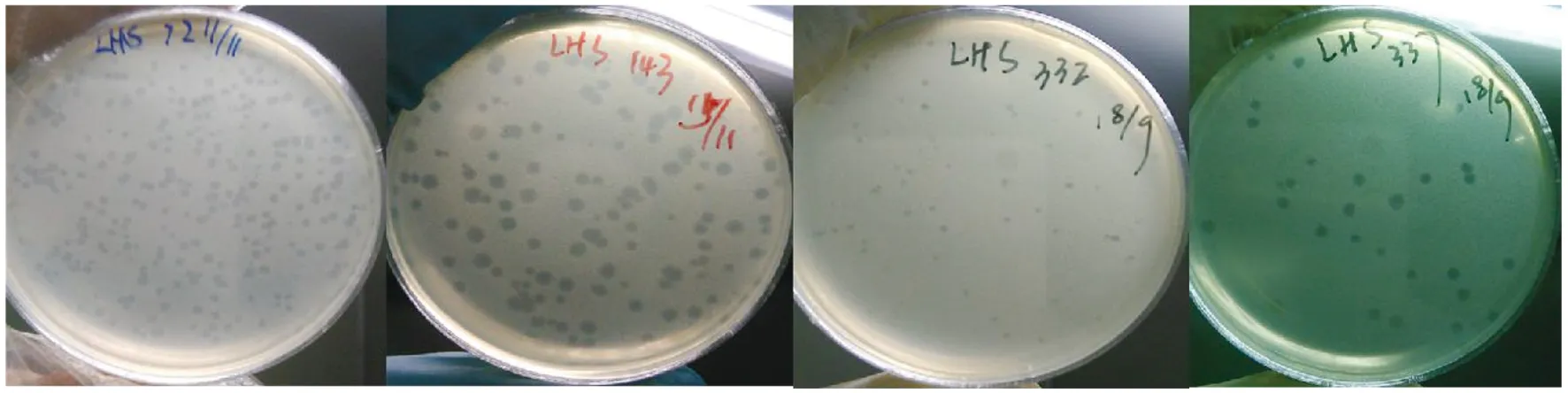
图2 梁河县鼠中分离4株鼠疫噬菌体的噬菌斑图Fig.2 Phage map of 4 Yersinia pestis phage isolated from rats in Lianghe County
2.4所分噬菌体的电镜照片 选取2株鼠疫噬菌体使用HT770透射电镜进行电镜扫描,放大200 000倍观察,如图3所示,这2株鼠疫噬菌体均为肌尾型鼠疫噬菌体,其中LHS143号鼠疫噬菌体头部直径大约71 nm,尾长129 nm,LHS250号鼠疫噬菌体头部直径大约50 nm,尾长86 nm。
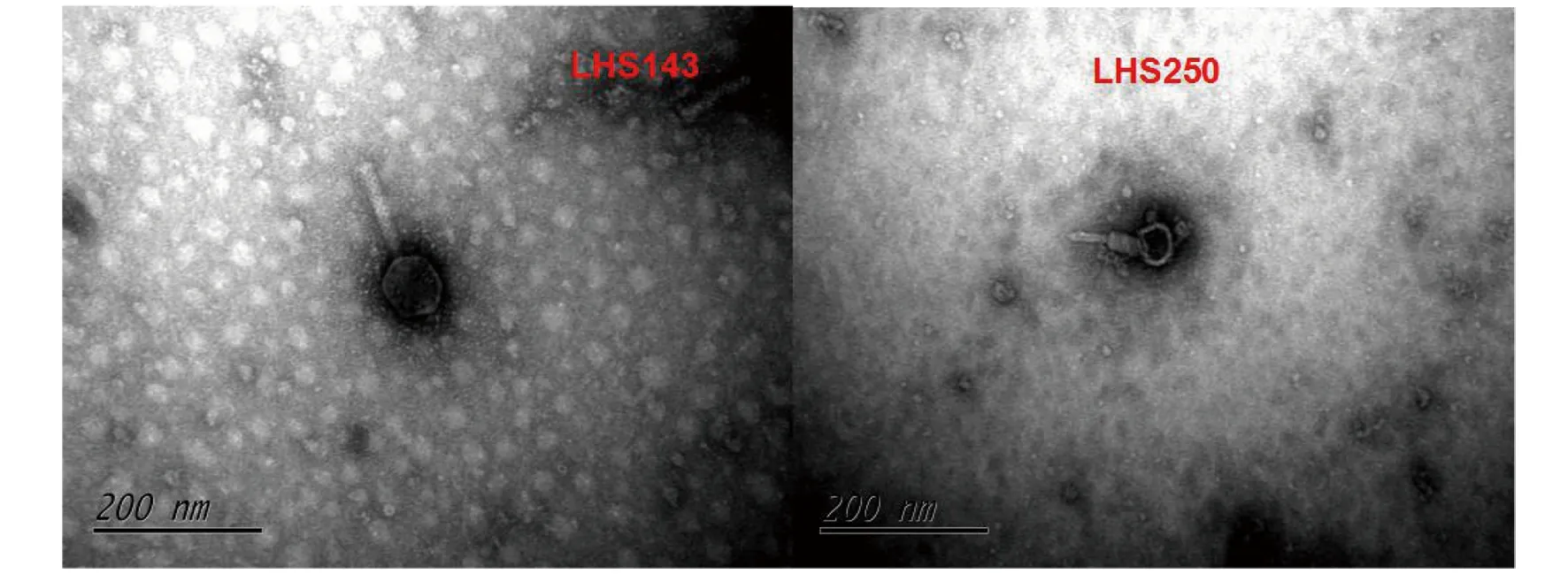
图3 梁河县鼠中分离2株鼠疫噬菌体的电镜照片Fig.3 Electron micrograph of 2 Yersinia pestis phage isolated from rats in Lianghe County
3 讨 论
3.1梁河家鼠鼠疫疫源地内广泛存在鼠疫噬菌体
梁河县在历史上曾猖獗地流行过鼠疫,自1955-1956年控制了人间和鼠间鼠疫以来,鼠间仍有疫情[7]。2001年发生一次人间腺鼠疫,之后进入静息期。本次研究在曾经流行过鼠疫的4个乡镇均分离到了鼠疫噬菌体,其中县城所在地遮岛镇分离到5株鼠疫噬菌体,分离率最高(16.13%),因其地理位置重要,人员密集,又是鼠疫流行过的疫点,加之鼠疫噬菌体分离率较高,在防控鼠疫中,应列在首位;芒东镇分离到12株鼠疫噬菌体,分离率最低(5.8%),证实了梁河家鼠鼠疫疫源地内广泛存在鼠疫噬菌体,并且其携带率高于剑川县与鹤庆县野鼠鼠疫疫源地鼠疫噬菌体携带率[5-6](分别是1.4%及0.65%)。鼠疫噬菌体主要依靠鼠疫菌而生存,样本中分离到鼠疫噬菌体,表明可能存在鼠疫菌或不久前曾感染过鼠疫菌[8]。本次研究在静息疫点分离到鼠疫噬菌体,而未分离到鼠疫菌,可能是因为鼠疫菌发生了某种用现有检测手段无法检出的变异,只能充当鼠疫噬菌体的宿主;也可能是因为鼠疫流行后期,在人为干预下逐渐建立了新的生态平衡,疫源地再次静息[9],从而未能分离到鼠疫菌;又或者因为鼠疫噬菌体较强的生存能力[10]致使其脱离宿主后还可生存较长时间,因此在相当长的静息期内仍可分离到鼠疫噬菌体。
3.2臭鼩鼱较黄胸鼠中有较高的鼠疫噬菌体携带率 梁河县家鼠鼠疫疫源地内存在十几种啮齿动物,其中以黄胸鼠为优势种,占59.83%,臭鼩鼱是仅次于黄胸鼠的鼠疫菌宿主[7],在鼠疫流行期,臭鼩鼱体内鼠疫菌的检出率明显高于黄胸鼠的检出率。本次研究从234只黄胸鼠中分离到19株鼠疫噬菌体,分离率为8.12%;从43只臭鼩鼱中分离到8株鼠疫噬菌体,分离率为18.6%,从11只锡金小鼠中分离到1株鼠疫噬菌体,分离率为9.1%;从仅有的1只卡氏小鼠中亦分离到1株鼠疫噬菌体。一个值得注意的现象是此次臭鼩鼱中鼠疫噬菌体的分离率要明显高于黄胸鼠中的分离率,分析其原因,可能是因为臭鼩鼱可栖息于室内、农耕区、灌木丛等地,活动范围与生存能力较黄胸鼠(家鼠)强,接触感染鼠疫噬菌体的机会更大。另外,在卡氏小鼠(家鼠)与锡金小鼠(家鼠)中均分离到鼠疫噬菌体,提示我们在鼠疫防控中,除了重视主要宿主黄胸鼠以外,其他家鼠也不能忽视。
3.3所分离的鼠疫噬菌体具有多态性 本研究中选取有代表性的2株噬菌体进行电镜扫描,LHS143与LHS250均为肌尾病毒科鼠疫噬菌体,NCBI中公布的已测序完成的鼠疫噬菌体中有3株属于肌尾病毒科噬菌体[11-12],且经基因比对3株之间无同源性[13],表明肌尾病毒科鼠疫噬菌体基因组具有多态性。本研究分离的噬菌体噬斑表现出多态性,推测它们的基因组也可能具有多态性,需要对其基因组进一步研究。
利益冲突:无
