奶牛乳脂性状候选基因EEF1D突变位点功能分析
2020-07-28蒋秋斐蔡正云黄增文冯小芳顾亚玲
蒋秋斐,蔡正云,黄增文,冯小芳,张 娟,顾亚玲,*
(1.宁夏大学 农学院,宁夏 银川 750021;2. 宁夏回族自治区畜牧工作站,宁夏 银川 750004)
奶牛业是世界公认的节能、经济、高效型产业。奶牛业发展水平是衡量一个国家发达程度和畜牧业现代化水平的重要标志。牛奶长久以来一直被人类视为最佳的膳食结构物质,其营养成分的合理性与全面性是其他任何食品都不可比拟的。为了保障我国奶质更加优质可靠,2017年我国建立了优质乳工程,并及时发布了优质乳核心标准,其中明确规定乳脂肪含量≥3.30%(美国≥3.20%)。然而,在实际生产中,增加牛乳中乳脂肪含量并不容易,面对如此严苛的要求,如何做到稳健提升奶质,是当前畜牧科学的重点研究方向之一。
真核翻译延伸因子1D(eukaryotic translation elongation factor 1 delta,EEF1D)是翻译延伸因子复合物(elongation factor 1,δeEF1)的eEFlBδ亚基。真核生物δeEF1由翻译延伸因子1A(elongation factor 1A,eEF1A)和翻译延伸因子1B(elongation factor 1B,eEF1B)组成,eEFlBδ是eEF1B中的催化亚基,发挥鸟嘌呤核苷酸转换因子的作用。Mulner-Lorillon等[1]提出EEF1D参与蛋白质合成;Gyenis等[2]发现其可能参与细胞周期活动;Delalande[3]在1998年发现EEF1D的两种可变剪切体;谢岩[4]研究发现,EEF1D有4种剪切体,并对EEF1D基因进行功能验证发现,在该基因上存在着一个与乳脂率显著相关的多态位点;还有一些研究表明,EEF1D可变剪切体在基因表达、进化分析、新陈代谢和生物复杂性等过程中发挥了非常重要的作用[5-8];Zhang等[9]在山羊中也发现EEF1D具有两种可变剪切体,其研究结果也同样表明其对乳脂合成具有极显著的相关性;刘轩[10]在中国荷斯坦牛EEF1D基因的启动子区和UTR区上检测到SNPs,通过关联性分析发现,其中2个位点与产奶量、乳脂率及乳蛋白率相关。
本课题组通过前期试验已将EEF1D基因确定为奶牛乳脂性状的候选基因,并通过预测发现,在其5′-UTR的G/A突变与乳脂率有极显著的关联。5′-UTR是mRNA中一段对翻译过程起调控作用的序列,主要通过其二级结构的蛋白质结合位点、核糖开关来调控翻译的起始。因而,本试验为了研究5′-UTR的G/A突变对奶牛乳脂含量的影响,主要通过转录因子结合位点预测及启动子活性分析,来检测该突变位点对目的基因表达的影响,分析突变位点对乳脂合成的调控作用,以期为奶牛育种提供理论依据。
1 材料与方法
1.1 细胞、载体及目的片段合成
Stbl3感受态细胞、293T细胞购自广州辉骏生物科技有限公司;pGL3-basic载体购自普洛麦格(北京)生物技术有限公司;EEF1Dwt/EEF1Dmut片段、引物分别由苏州泓迅生物科技有限公司和广州艾基生物科技有限公司合成。
1.2 主要试剂
限制性内切酶、T4 DNA连接酶、Lipofectamine 2000转染试剂购自赛默飞世尔科技(中国)有限公司;DNA回收试剂盒、普通质粒小提试剂盒I型购自中美泰和生物技术有限公司;PrimeSTAR Max Premix(2×)、TaKaRaTaqMax购自宝生物(大连)有限公司;DMEM培养基、PRMI 1640、南美胎牛血清FSP500、胰酶、氯化钠(NaCl)、Dual-Luciferase®Reporter Assay System(E1910)购自美国Invitrogen公司。
1.3 试验方法
1.3.1 转录因子结合位点的分析与合成
通过在线软件ALGGEN(http://alggen.lsi.upc.es/)、TRANSFAC(http://www.generegulation. com/pub/databases.html#transfac)对奶牛EEF1D基因5′非翻译区突变位点前后的转录因子进行预测后,根据ENSBTAG00000014643序列号、NCBI AC_000171.1确定奶牛EEF1D基因的启动子区,选取目的片段大小为981 bp,并由苏州泓迅生物科技有限公司合成EEF1D基因的野生型(EEF1Dwt)和突变型(EEF1Dmut)片段。
1.3.2 重组表达载体(pGL3-basic-EEF1D-wt/mut)的构建和鉴定
将目的基因用KpnⅠ和HindⅢ双酶切后连接至经相同酶切的pGL3-basic载体中,转化至Stbl3感受态细胞。挑取单克隆,经PCR鉴定和抽提质粒用KpnⅠ和HindⅢ双酶切鉴定,获得阳性克隆进行测序。
1.3.3 双荧光素酶报告基因实验
将构建好的重组质粒pGL3-basic-EEF1D-wt/mut和空载体质粒pGL3-basic纯化回收后,分别与转染试剂(脂质体2000)按一定比例混匀,在37 ℃含5%的CO2培养箱中,与293T细胞共同孵育。48 h后,收集细胞进行双荧光素酶活性实验,以检测目的基因启动子活性,通过检测其萤火虫荧光素酶发光值F和加入STOP&GLO Reagent后测得对应海肾荧光素酶发光值R。计算出F和R的比值(即Luc),来反映EEF1D基因不同启动子片段的活性。
1.4 统计学分析
采用统计分析软件GraphPad Prism 5进行分析,采用t检验进行差异性检验。
2 结果与分析
2.1 转录因子结合位点的预测
通过在线软件对候选基因5′-UTR突变位点前后的转录因子结合位点进行预测,预测阈值最大矩阵相似率设定为87%,发现c.-488 A>G发生G→A突变前后,引起转录因子Sp3(specificity protein 3)转录起始结合位点位置改变,改变前后位点之间相差33 bp。
2.2 启动子活性分析
2.2.1 重组质粒琼脂糖凝胶检测结果
重组质粒经KpnⅠ和HindⅢ双酶切后,分别得到981、1 500 bp的片段,凝胶电泳检测结果如图1,观察到的条带与预期位置相符。根据克隆测序结果显示,测序结果与预期序列100%匹配。
2.2.2 连接产物测序验证
对重组质粒pGL3-Basic-EEF1Dwt/pGL3-Basic-EEF1Dmut分别进行测序验证(图2),经分析,插入片段长度均为981 bp,且序列与预期序列完全一致,表明重组载体构建成功。
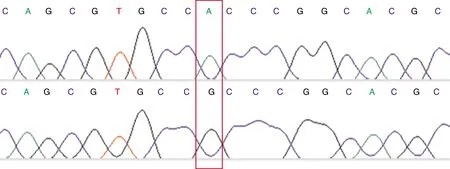
图2 测序结果比对Fig.2 The results of sequencing comparison

M,DL2000 marker;1,经KpnⅠ和HindⅢ酶切pGL3-Basic-EEF1Dwt的载体;2,经KpnⅠ和HindⅢ酶切pGL3-Basic-EEF1Dmut的载体。M,DL200 marker; 1,The vector for pGL3-Basic-EEF1Dwt by KpnⅠ and HindⅢ enzyme digestion; 2,The vector for pGL3-Basic-EEF1Dmut by KpnⅠ and HindⅢ enzyme digestion.图1 酶切电泳结果Fig.1 The electropherogram of production after enzyme digestion
2.3 EEF1D基因5′非翻译区突变的启动子活性分析
双荧光素酶活性检测实验结果见图3,根据结果显示,pGL3-EEF1Dwt、pGL3-EEF1Dmut的双荧光素酶活性相对于basic显著升高(P<0.01),EEF1Dmut的活性大约是EEF1Dwt的3倍(P<0.01),说明突变位点促进目的基因的表达。结合转录因子结合位点的预测结果,可以推测c.-488A>G突变位点是影响EEF1D基因表达的关键突变位点。
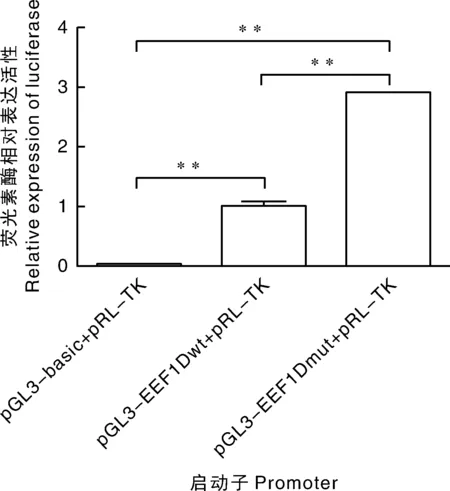
*表示P<0.05;**表示P<0.01。* represented P<0.05; ** represented P<0.01.图3 野生型与突变型重组载体的启动子活性分析Fig.3 Promoter activity analysis of wild type and mutation type recombination vector
3 讨论
课题组前期发现EEF1D基因与乳脂率极显著相关,并将此基因作为奶牛乳脂性状的功能候选基因。对于该基因,前期的研究主要集中在EEF1D可变剪切体方面[5],发现EEF1Da是EEF1D在泌乳期奶牛乳腺组织中的存在形式,其他有关EEF1D在乳脂合成通路中的作用和功能方式的研究还鲜有报道。本研究发现,EEF1D基因的5′非翻译区存在一个突变位点,通过进一步经转录因子预测结合启动子活性分析发现该突变会引起EEF1D基因的表达量表达上调,最终导致转录因子Sp3转录起始位点后移33 bp。
SNP和甲基化都会影响基因的表达。SNP是指单个核苷酸的变异导致DNA序列多态性,可以位于DNA序列的任何位置,包括单个碱基转换、颠换、缺失和插入,前两者在实际情况中较常发生,发生概率比是2∶1。后两种情况很少见。SNP在GC序列出现最为常见,且多见于C/T的转换[11]。在自然条件下,核苷酸发生突变的频率并不高,或者说很低,而SNP只有在单碱基发生突变后才会产生,所以SNP与其他标记相比,遗传稳定性要更高[12-16],而且SNP会导致个体发生变异或病变,基于这两点检测SNP可以为研究个体性状的遗传机制提供依据[17]。甲基化是指从SAM等活性甲基化合物上将-CH3转移到其他类型化合物的过程,包括DNA和组蛋白两种甲基化。DNA甲基化常见发生于CpG位点,蛋白质甲基化常见发生于序列中Lys或Arg,CpG岛是指GC含量大于50%,长度大于200 bp的区域,在基因的启动子和外显子区域较常见。刘轩[10]在EEF1D基因启动子区和UTR区均检测到了CpG岛的存在,并进行甲基化分析,发现乳腺组织中的甲基化水平显著低于其他组织,说明EEF1D基因启动子区的CpG岛的甲基化程度直接影响EEF1D基因的表达。由于EEF1D基因启动子目的序列的GC含量高于90%,再结合前人研究结果和序列的特殊性,本研究最终通过化学合成方法获得了目的序列。同时,因为表观遗传学修饰侧重于研究的是同一基因型而表型不同的现象,所以本研究未对CpG岛甲基化程度等方面开展研究。
转录因子可以按其基本结构、锌指结构、HTH划分为不同的类型。Sp3是含有3个锌指结构的因子,具有转录激活或者转录抑制的双重作用,但在大多数情况下,其发挥转录抑制的作用。Sp3是Sp(specificity protein,Sp)/Kruppel样(Kruppellikefactors,KLF)蛋白家族的成员之一,该家族多数成员与脂肪细胞分化的调控有关。等级越高的物种,具有Sp/KLF蛋白的种类也越多,从NCBI数据库查询可知,在果蝇上发现了3种Sp/KLF蛋白,在人体内预测存在有25种,己经克隆出了24种(Sp9尚未被克隆)[18]。
通过在线软件进行EEF1D野生型和突变型转录因子结合位点预测,发现有351个存在的可能结合的转录因子。其中,除了Sp3转录起始位点预测结果变化较大,其他均未发生变化。结合启动子活性分析实验,EEF1D突变型的活性大约是EEF1D野生型的3倍。这样可能是因为Sp3转录起始位点的改变,增强了其与家族基因或与目的基因或特定序列的结合强度,从而促进了目的基因的表达。Sp3转录起始位点变化对奶牛奶质中乳脂含量的调控机制,还有待进一步深入的研究。
