天津海域鱼卵、仔稚鱼资源动态研究
2020-07-22谷德贤王婷徐海龙王娜吴宁李文雯刘国山
谷德贤,王婷,徐海龙,王娜,吴宁,李文雯,刘国山
(天津市水产研究所,天津 300221)
天津海域地处渤海湾底部,有蓟运河、潮白新河、永定新河、海河、大沽河、独流减河、青静黄排水渠、子牙新河、北排河等多条河流注入,是花鲈、小黄鱼、鳀等多种鱼类的主要产卵场之一。鱼类种群的数量变化主要受补充量的影响,种群的补充机制决定了渔业资源的世代发生量和生物资源的可持续产量[1]。目前,对鱼卵、仔稚鱼的研究多集中在长江口及邻近水域[2-5],主要探讨了鱼卵、仔稚鱼的丰度变化,以及鱼卵、仔稚鱼与温度、盐度、入海径流、浮游动物的关系等。此外,还有针对不同海区的鱼卵、仔稚鱼研究,如万瑞景等[6-8]研究了黄海和东海的鱼卵、仔稚鱼分布,渤海的相关研究主要集中在莱州湾[9-12],于旭光等[13]研究了辽东湾的鱼卵、仔稚鱼组成,李宝华等[14]统计了渤海湾1999—2002年的鱼卵数量。鱼卵、仔稚鱼的存活率及数量是鱼类资源进行补充和可持续利用的基础[7-8],但鱼卵、仔稚鱼阶段也是鱼类生活史中最脆弱的阶段,较小的环境变化也可能对资源的补充造成较大影响。近年来,由于大规模围海造地占据了大量的滨海湿地和浅海资源[15]、入海径流量变化、近海富营养化导致的赤潮和水母暴发等生态灾害,以及环境污染、海洋开发、渔业捕捞、气候变化等原因造成的鱼类群落结构发生较大的变化,使得渤海的产卵场功能不断退化[16]。
20世纪80年代,天津市开展的海岸带和海涂资源综合调查中的浮性鱼卵和仔稚鱼调查[17],较为系统地描述了天津近岸海域鱼卵、仔稚鱼的种类组成、时空分布、经济鱼类的产卵时间等,此后的研究较少[14,18]。而针对整个渤海的鱼卵、仔稚鱼研究中[5],布设的站位极少涉及天津海域。为此,本研究中通过分析2012—2018年天津海域鱼卵、仔稚鱼及环境调查数据,分析鱼卵、仔稚鱼的时空分布变化特征,旨在为天津海域渔业资源补充群体的研究提供参考。
1 材料与方法
1.1 数据来源
数据来源于天津市水产研究所2012—2018年进行的29个航次367个站次对鱼卵、仔稚鱼及相关环境因子的调查。具体调查航次为2012年6、8、9、11月,2013年5、6、8、10月,2014年5、6、7、8、11月,2015年2、5、6、7月,2016年6、7、8、10月,2017、2018年的5、6月上旬、6月下旬、7月。调查范围为天津海域(38°25′~39°20′N、117°30′~118°20′E),调查船主机功率为176 kW,船长为24.88 m,型宽为5 m,型深为2.2 m,总吨位为64 t。
1.2 方法
1.2.1 样品的采集与鉴定 鱼卵、仔稚鱼调查使用大型浮游生物网(网长280 cm,网口内径80 cm,网口面积0.5 m2)在海水表层进行水平拖网10 min,船速为1~2 n mile/h,网口系有网口流量计。采集到的样品用5%的甲醛溶液固定,带回实验室对鱼卵、仔稚鱼标本进行分拣、种类鉴定、个体计数等分析。相关环境因子如温度、盐度、溶解氧、营养盐、叶绿素a、浮游生物等参照《海洋调查规范》(GB12763—2007)[19]的相关要求进行。
1.2.2 鱼卵、仔稚鱼丰度 丰度计算公式为
Ga=Na/(S×L×C)。
(1)
其中:Ga为单位体积海水中鱼卵或仔稚鱼个体数(ind./m3);Na为全网鱼卵或仔稚鱼个体数(ind.);S为网口面积(m2);L为流量计转数;C为流量计校正值。
1.2.3 优势种和重要种 优势度计算公式为
IRI=N×F。
(2)
其中:IRI为优势度;N为每航次调查过程中某种鱼卵或仔稚鱼个体数量占鱼卵或仔稚鱼总量的比例(%);F为每航次调查过程中某种鱼卵或仔稚鱼出现的频率百分比(%)。参考卞晓东等[16]对鱼卵、仔稚鱼优势种和重要种的判断标准:IRI>1000为优势种;鱼卵IRI=500~1000为重要种,仔稚鱼IRI=350~1000为重要种;鱼卵IRI=50~500为一般种,仔稚鱼IRI=50~350为一般种;IRI=10~50为少见种;IRI<10为稀有种。
1.2.4 环境因子相关性分析 应用广义加性模型(Generalized additive model,GAM)分析鱼卵、仔稚鱼丰度与影响因素之间的关系,模型表示为
(3)
其中:Y为鱼卵或仔稚鱼或鱼卵与仔稚鱼总体丰度;g(Y)为连接函数;xi为各环境因子;fi(xi)为g(Y)与第i个环境因子的非参数函数;k为环境因子个数;ε为误差项。
8月29日,智利化学矿业公司(SQM)市场发展部副总裁Alfredo Doberti一行到访中农控股,中农控股副总经理王蓓会见了来访客人,双方就智利氮钾二元肥以及全水溶肥料的合作事宜进行了深入交流。
本研究中,连接函数为自然对数,模型采用样条平滑法,分布函数族为泊松分布,模型的误差分布估计为高斯分布,具体计算过程参见谷德贤[20-21]中的方法,分析通过R 3.5.2 软件中mgcv包实现。
2 结果与分析
2.1 种类组成
天津海域7年共监测到鱼卵、仔稚鱼31种,包括鱼卵17种、仔稚鱼23种,隶属于7目16科31种,其中1种鉴定到科,29种鉴定到种,未定种1种。鱼卵的种类组成情况如下:2012—2018的调查主要集中在5月—8月,鱼卵种类数最多的月份集中在6月,最多时为12种,主要为斑鰶Konosiruspunctatus、短吻红舌鳎Cynoglossusjoyneri、青鳞小沙丁鱼Sardinellazunasi、石首鱼科Sciaenidae、赤鼻棱鳀Thryssakammalensis等;9月调查了1个船次,采集到的1种鱼卵;11月和2月各调查了1个航次,均未采集到鱼卵;10月调查了3个航次,1个航次(2013年)未采集到鱼卵,另外2个航次采集到的鱼卵均为花鲈Lateolabraxjaponicus;鱼卵种类数随年份总体上呈现增加的趋势,其中,2018年采集的鱼卵种类数最多,为14种,2013年种类最少,为5种(表1)。
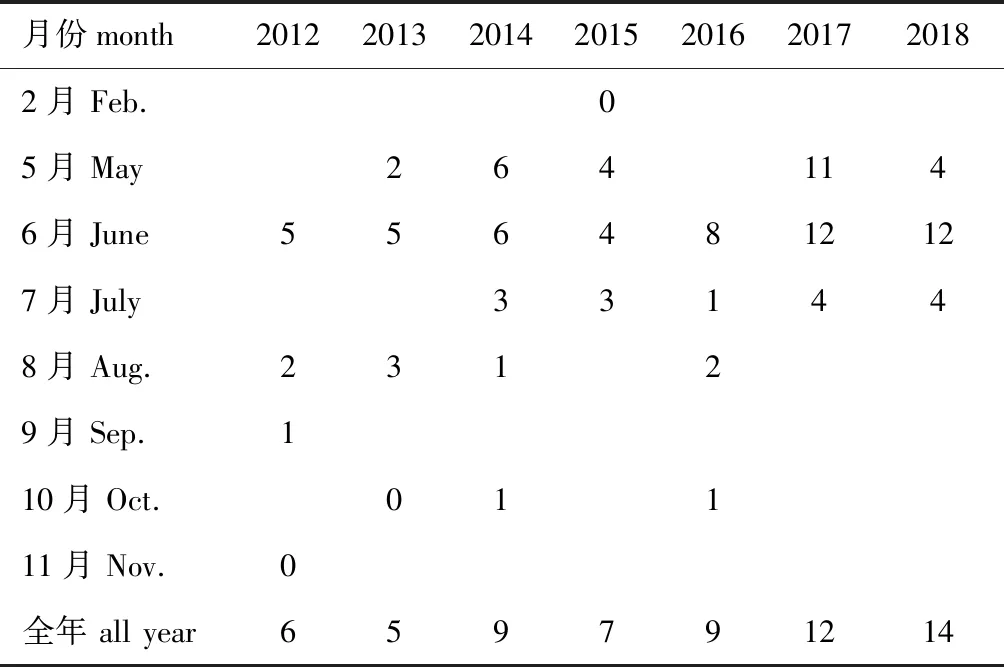
表1 鱼卵种类数的变化Tab.1 Changes in fish species number of the eggs
仔稚鱼的种类组成情况如下:5—8月均有仔稚鱼出现,6月仔稚鱼种类最多,最多时为8种,主要为斑鰶、赤鼻棱鳀、鮻Planilizahaematocheila、黄鲫Setipinnatenuifilis、矛尾虾虎鱼Chaeturichthysstigmatias等;9月调查了1个航次,未采集到仔稚鱼,10月调查了3个航次,有2个航次未采集到仔稚鱼,1个航次采集到了赤鼻棱鳀和花鲈的仔稚鱼;11月和2月各调查了1个航次,分别采集到2种(黄鲫和花鲈)和1种(玉筋鱼Ammodytespersonatus);各年间仔稚鱼的种类数波动较小,2018年种类最多,为12种,2013年种类最少,为7种(表2)。
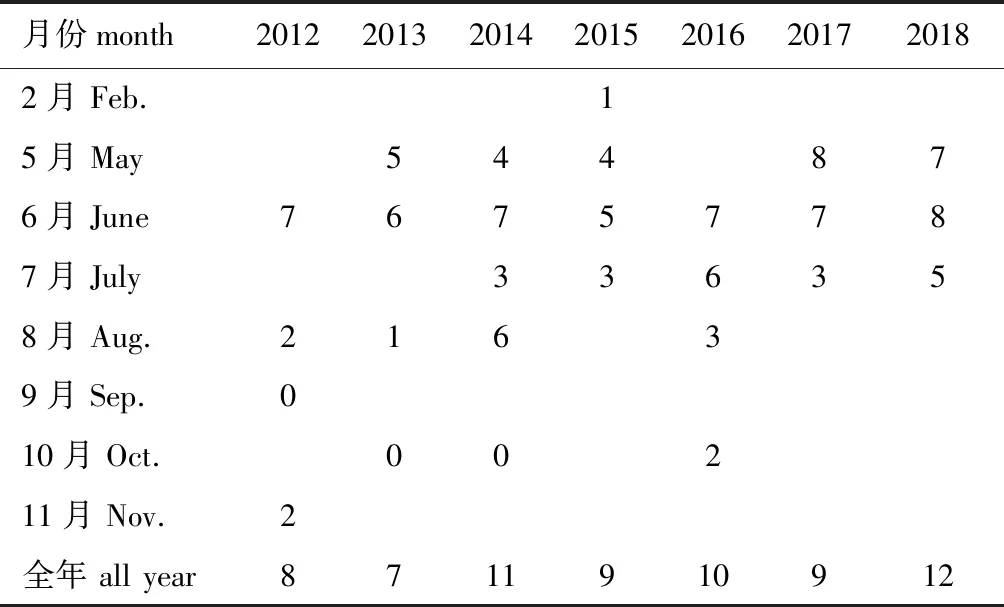
表2 仔稚鱼种类数的变化Tab.2 Changes in fish species number of the larvae
2.2 数量分布
2012—2018年的调查集中在5月—8月,鱼卵丰度为1~5 ind./m3的月份出现在2014、2016年的6月及2017年的5月和6月(表3)。仔稚鱼丰度大于5 ind./m3的月份出现在2012年6月,丰度为1~5 ind./m3的月份出现在2015年5月(表4)。
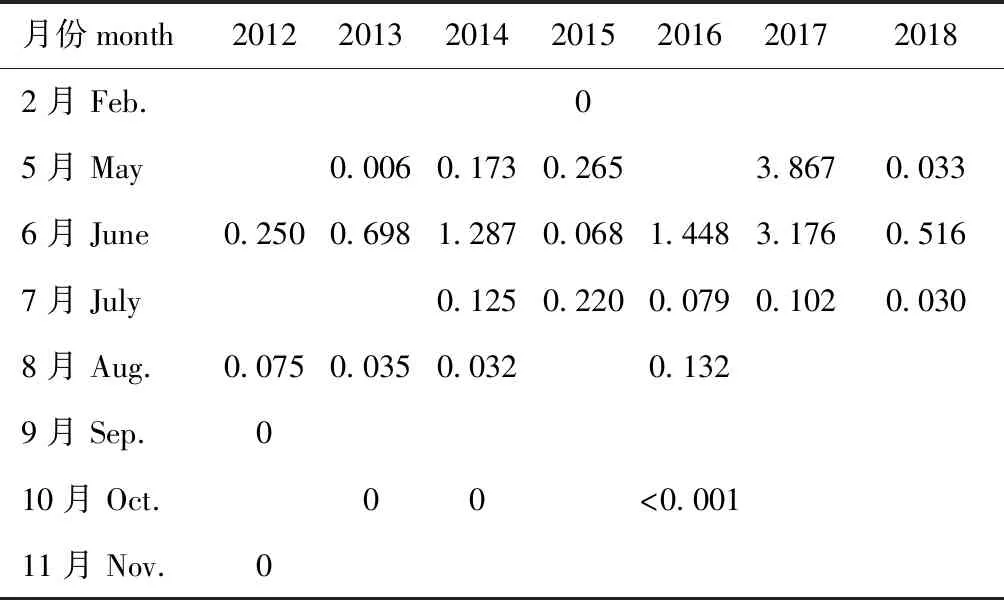
表3 鱼卵丰度的变化
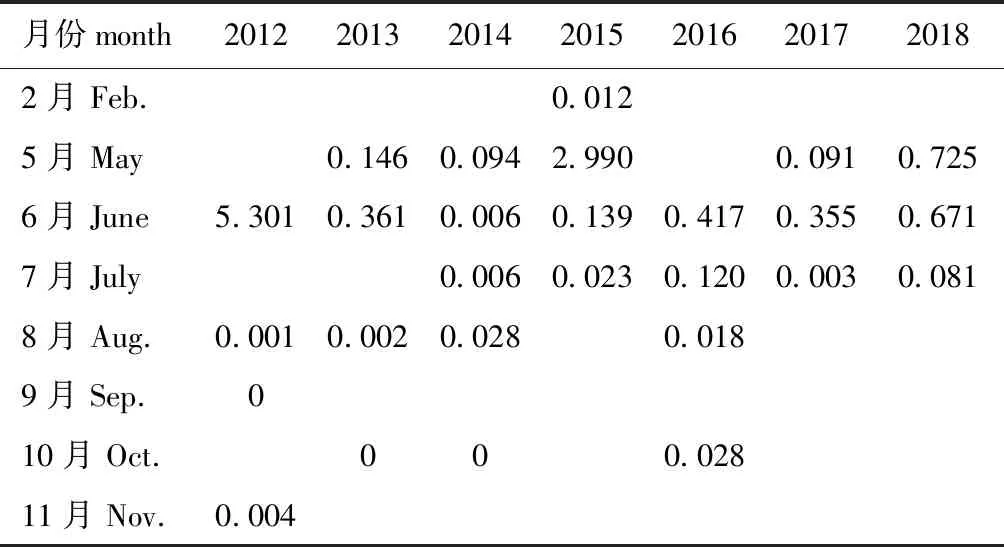
表4 仔稚鱼丰度的变化
2.3 优势种
从如表5可见:斑鰶鱼卵主要为5月的优势种;石首鱼科鱼卵主要为5月—7月的优势种和重要种;短吻红舌鳎鱼卵主要为6月—8月的优势种;花鲈鱼卵为10月的重要种。从表6可见,玉筋鱼仔稚鱼为2月的优势种;鮻仔稚鱼主要为5月的优势种;赤鼻棱鳀仔稚鱼主要为6月—8月的优势种;斑鰶仔稚鱼主要为5月—6月的优势种;花鲈仔稚鱼为10月—11月的优势种。
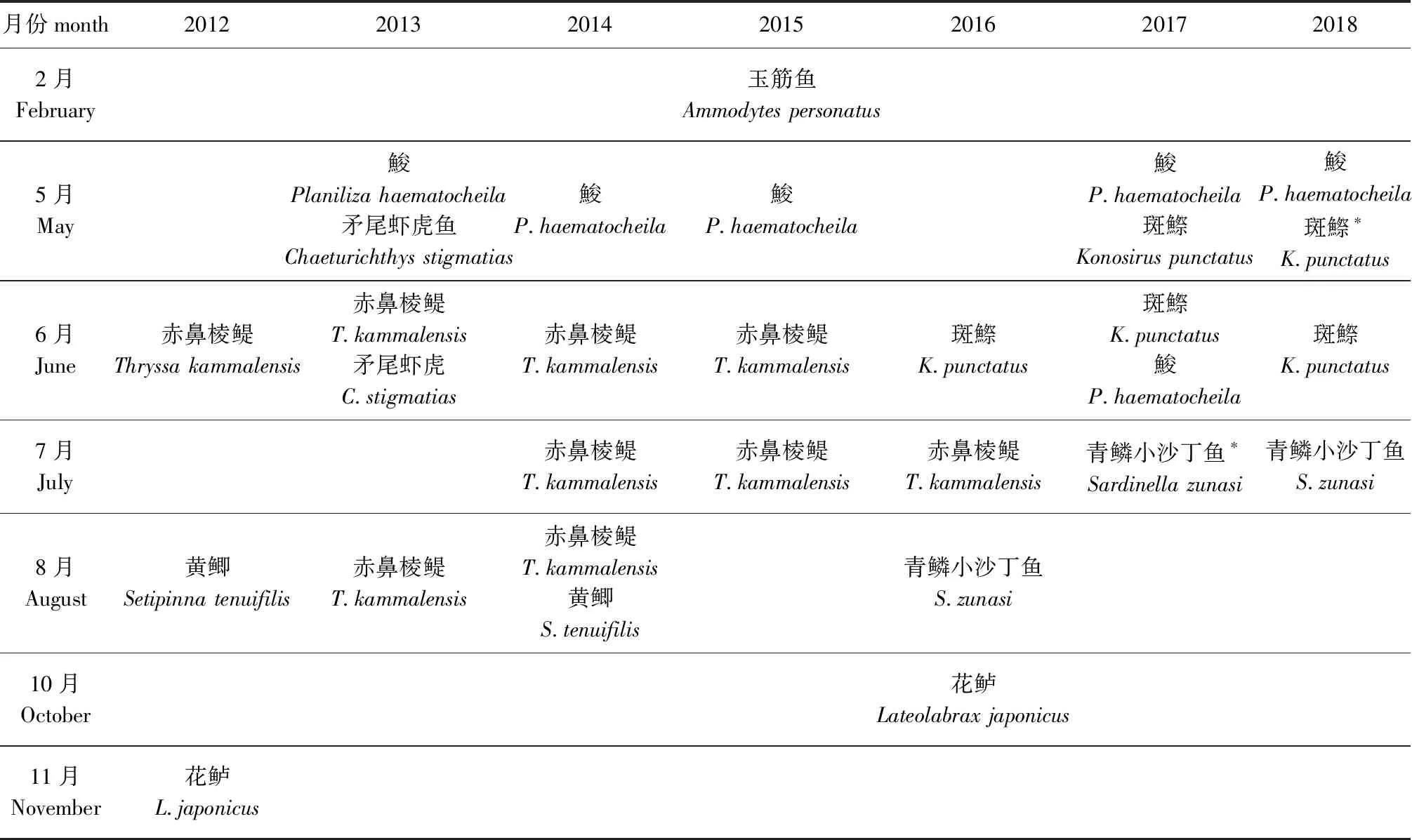
表6 仔稚鱼优势种和重要种Tab.6 Dominant species and important species of fish larvae
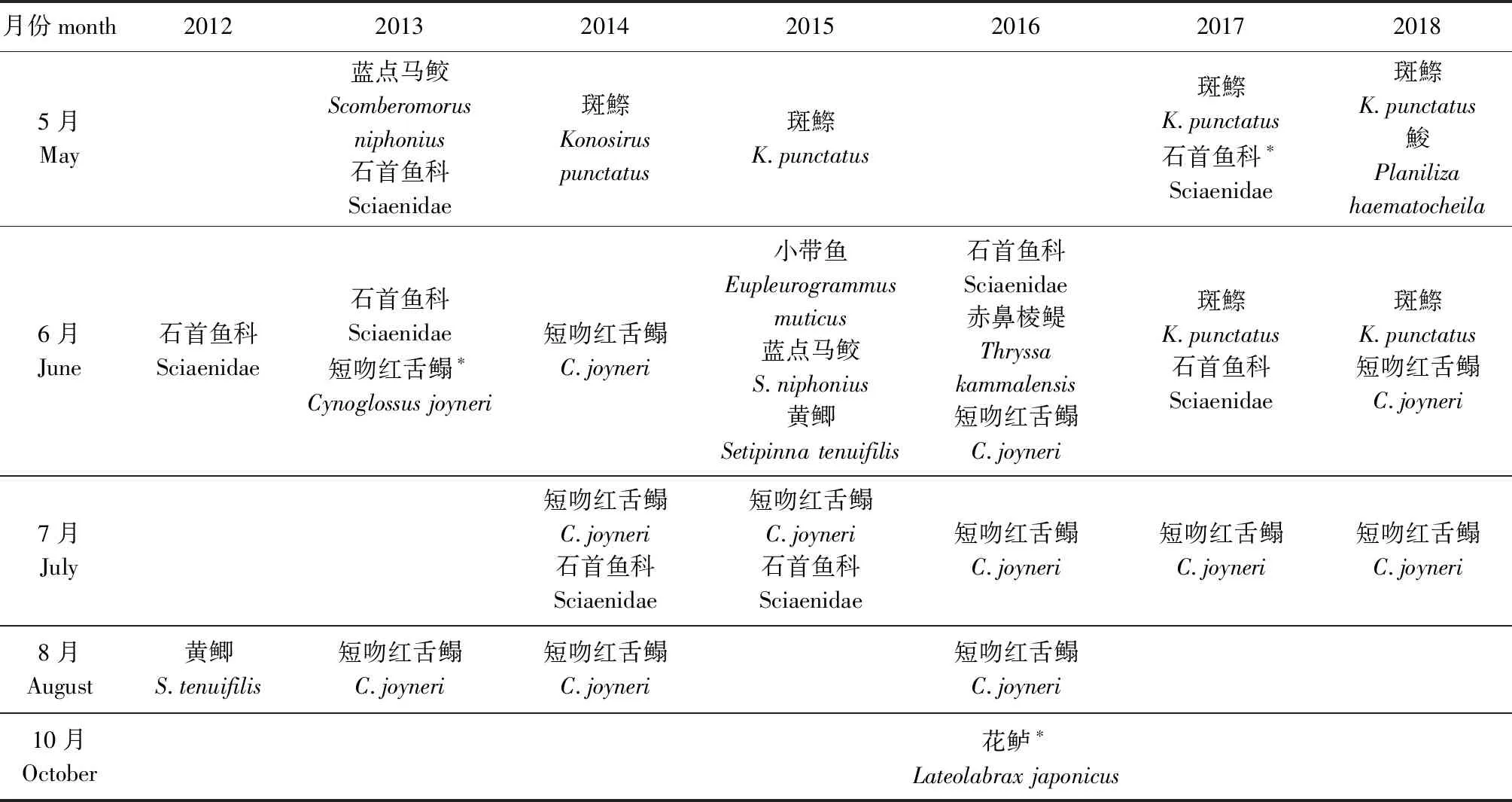
表5 鱼卵优势种和重要种Tab.5 Dominant species and important species of fish eggs
2.4 空间分布
2012—2018年鱼类早期补充群体的丰度分布如图1所示。2012年鱼类早期补充群体(鱼卵和仔稚鱼),6月主要分布在南部近岸海域,其次为曹妃甸外海流进天津海域处;8月主要分布在北部海域,总体量较少。2013年鱼类早期补充群体,5月主要分布在近岸;6月向中间水域、外海水域移动;8月分布在中部塘沽海域。2014年鱼类早期补充群体,5月主要分布在南部外海及北部近岸;6月由远海向近岸增多,由南部海域向北增多;8月鱼类早期补充群体均匀分布,北部较多(2014和2015年的图中缺少7月航次数据,因为7月航次调查的站位与其他月份调查站位不一致,故只做了一个断面的调查图)。2015年鱼类早期补充群体,5月近岸多于外海,南部海域多于北部,而后向外海移动;7月近岸和外海鱼卵多,中间水域仔稚鱼多,形成内外交混的格局。2016年鱼类早期补充群体,6月主要分布在北部海域的南侧和南部海域的中部;7月南部海域和北部海域群体向中部海域移动;8月天津海域近岸及外海较多,中间水域较少。2017年鱼类早期补充群体,5月外海多于近岸,南部海域多于北部;6月鱼卵增多,南部海域多于北部,并逐渐形成外海和近岸多、中间海域少的分布格局;7月南部海域多于北部海域,分布较均匀。2018年鱼类早期补充群体,5月分布同2017年5月;6月鱼卵增多,南部海域多于北部,并逐渐向外海移动;7月南部海域多于北部海域,分布较均匀。
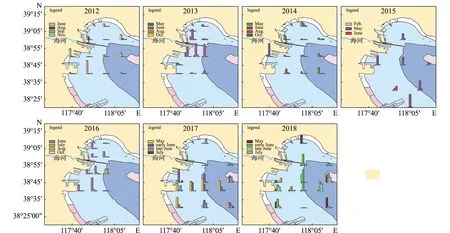
图1 2012—2018年鱼类早期补充群体的丰度分布图(2014、2015年7月数据未显示)Fig.1 Abundance of early fish cruitments in 2012—2018(Data for July 2014 and July 2015 are not shown in the figure)
2.5 主要环境影响因子
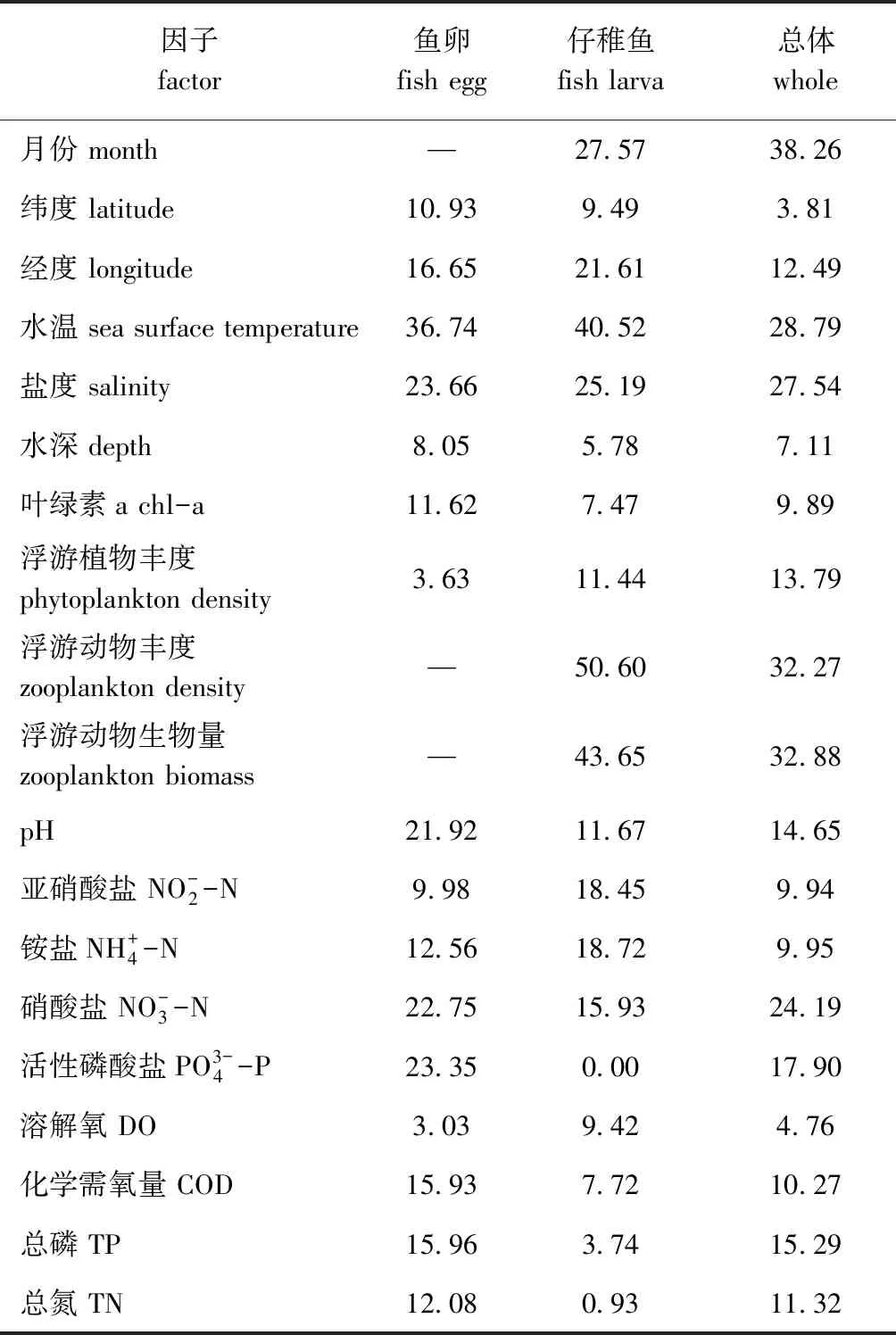
表7 鱼卵、仔稚鱼丰度与相关因子GAM的解释偏差
3 讨论
3.1 种类及优势种变化
对比1983年天津市海岸带和海涂资源综合调查中的鱼卵、仔稚鱼结果[6],近年来减少的种类有鳓Ilishaelongata、安氏新银鱼Neosalanxanderssoni、银姑鱼(白姑鱼)Pennahiaargentata、黄姑鱼Nibeaalbiflora、牙鲆Paralichthysolivaceus、半滑舌鳎Cynoglossussemilaevis、淞江鲈Trachidermusfasciatus、褐菖鲉Sebastiscusmarmoratus、长绵鳚Zoarceselongatus、红娘鱼Lepidotriglamicroptera,增加的种类有鱼銜Callionymushuguenini、矛尾复虾虎、髭缟虾虎鱼Tridentigerbarbatus、六丝钝尾虾虎鱼Amblychaeturichthyshexanema、许氏平鲉Sebastesschlegelii(近年增殖放流种)等。据张闰生等[17]的研究结果,1983年鱼卵、仔稚鱼优势种为斑鰶、青鳞小沙丁鱼、鳀Engraulisjaponicus、赤鼻棱鳀和叫姑鱼Johniusgrypotus,其中以鳀的数量最多。据本研究中2012—2018年对鱼卵、仔稚鱼的调查结果,斑鰶依然为优势种,鳀和叫姑鱼不再是优势种,曾经数量最多的鳀已经非常少。
3.2 丰度变化
天津海域1999—2002年的鱼卵、仔稚鱼丰度[14]分别为6.05、2.67、2.40、6.0 ind./m3,2008年的丰度[18]为2.88 ind./m3,而2012—2018年的平均丰度为0.809 ind./m3,较2008年降低了71.9%,较1999—2002年平均值(4.28 ind./m3)降低了81.1%。若航速为2 kn,拖网时间为10 min,大型浮游生物网的网口面积为0.5 m2,则对于10 min的拖网而言,1 ind./m3鱼卵、仔稚鱼约为309 ind./net。相应地,2012—2018年5月和6月鱼卵数量的最大值分别为1195粒和982粒,而1983年调查[17]的相应结果(拖网5 min)分别为127 077粒和10 857粒,5月和6月鱼卵的最大数量约为1983年相应月份的0.94%和9.04%。2012—2018年5月和6月的仔稚鱼平均数量分别为924尾和2168尾,1983年调查的相应结果分别为79 837尾和14 077尾,5月和6月仔稚鱼的最大数量约为1983年相应月份的1.16%和15.40%。这说明,5月的鱼卵、仔稚鱼的数量衰退程度较6月更严重。
3.3 季节分布
1983年调查结果显示,5月的鱼卵、仔稚鱼数量占全年(不含1月、2月和12月)数量的86.5%,6月占10.4%[17]。2012—2018年(表3和表4)调查显示:各月的鱼卵、仔稚鱼丰度存在年际间差异,2012—2016年和2018年,6月的丰度较大;2017年,5月和6月的丰度差别较小;2016—2018年,6月的调查分为月初和月末2次,6月末的丰度一般低于6月初的丰度。由此可见,这7年的鱼类产卵的高峰期可能在5月到6月初。与1983年的调查结果相比,产卵的高峰期略向后移。杨艳艳等[12]于2014、2015年的5月和8月对莱州湾的调查表明,5月采集的鱼卵、仔稚鱼占了绝大部分。于旭光等[13]于2008年5月、7月、9月和10月对辽东湾的鱼卵、仔稚鱼的调查表明,5月和7月是主要产卵季节。这说明,5月可能为渤海三个海湾共有的产卵盛期。
3.4 环境影响因子
王宏田等[22]从养殖的角度分析了影响鱼卵孵化和仔鱼发育的主要影响因子,主要为盐度、光照、溶解氧、温度、pH等。本研究中选取了月份、经纬度、水温、盐度、水深、叶绿素a、浮游植物(丰度)、浮游动物(丰度和生物量)、溶氧、pH、硝酸盐氮、亚硝酸盐氮、氨氮、磷酸盐、总氮、总磷、化学耗氧量等因素。基于GAM分析,月份、浮游动物、温度是影响鱼类早期补充群体的主要因素,然后是盐度因子。月份和温度两个因素是密切相关的,温度是影响鱼类早期补充群体的主要影响因子[23]。浮游动物是鱼类早期补充群体的重要食物来源[3],同时也有某些浮游动物摄食鱼卵、仔稚鱼[24],浮游动物决定着鱼类早期补充群体的生存生长,是其重要的生物环境影响因子。本研究中发现,天津海域鱼卵、仔稚鱼对温度的敏感性高于盐度,杨艳艳等[12]对莱州湾鱼卵、仔稚鱼的研究也有相似发现。此外,鱼卵无游泳能力,仔稚鱼的游泳能力弱,海流也是重要的影响因素。据山广林[25]报道,夏季天津附近海域的表层余流沿海岸向北,在大沽灯塔附近向东流或东南偏南流,流速一般小于等于10 cm/s;毕聪聪[26]对渤海四季环流的模拟中,表层余流的情况也大致相同,环流的流速大多小于等于5 cm/s,远小于潮流流速,最大流速约30 cm/s。海流可以改变鱼卵、仔稚鱼的空间分布,其影响范围、程度等有待进一步研究。
根据本研究中2012—2018年的调查结果,5月和6月为天津海的产卵盛期,5月鱼卵的优势种为斑鰶,仔稚鱼的优势种为鮻;6月鱼卵的优势种为石首鱼科、斑鰶和短吻红舌鳎,仔稚鱼的优势种为赤鼻棱鳀和斑鰶。鱼卵、仔稚鱼在6月份平均生物丰度最大,分别为0.969、0.600 ind./m3;5月份生物丰度略低于6月,分别为0.922、0.586 ind./m3。鱼卵5月和6月的数量约为1983年相应月份的0.11%和1.38%,仔稚鱼5月和6月的数量约为1983年相应月份的0.11%和0.66%。与20世纪80年代相比,天津的海岸线已发生了巨大变化,人类活动对海洋的影响加剧,天津海域的鱼类早期补充群体在种类、优势种、数量等方面,较历史调查结果均发生了明显的变化,传统的经济种类减少明显,鱼卵、仔稚鱼数量下降。
3.5 天津海域鱼卵、仔稚鱼保护建议
2018年之前,渤海从6月1日进入禁渔期,2018年后,渤海从5月1日进入禁渔期。2018年的鱼卵、仔稚鱼丰度虽有增加但未明显增加(表3和表4)。为了更好地发挥禁渔的作用,保护和利用鱼类资源,建议如下:
(1)将鱼卵、仔稚鱼的调查纳入常规监测,并监测产卵群体的分布情况。
(2)在产卵的关键时期增加调查频次,如5月—7月,至少每半个月调查一次。
(3)鱼卵统计应区分活卵和死卵,以更好地了解鱼卵的存活情况。
(4)多采用声学、原位观察等不损害资源生物的技术,把握产卵群体和鱼卵、仔稚鱼的整体动态变化。
