基于16S rRNA与COI基因40种石斑鱼亚科鱼类分子系统进化关系
2020-07-21梁日深陈铭廖国威张卓为张癸新陈轶之林蠡
梁日深,陈铭,廖国威,张卓为,张癸新,陈轶之,林蠡*
( 1. 仲恺农业工程学院 动物科技学院,广东 广州 510225;2. 广东省水环境与水产品安全工程技术研究中心,广东 广州510225;3. 广州市水产病害与水禽养殖重点实验室,广东 广州 510225;4. 深圳市里维生鲜科技开发有限公司,广东 深圳518052)
1 引言
石斑鱼亚科(Epinephelinae)鱼类隶属鲈形目(Perciformes)、鲈亚目(Percoidei)、鮨科(Serranidae),广泛分布于热带、亚热带大陆沿海及岛礁海域,是重要的海洋经济食用与观赏鱼类。世界上石斑鱼亚科约15属150多种[1],我国主要有10属50多种,大部分分布于东海及南海[2-5]。
石斑鱼种类繁多,形态分类主要依据体型,鳍的结构与数目,体色与斑纹等有限的外部形状进行鉴定。而石斑鱼近缘物种十分相似,许多种类在外部形态特征如体色、斑纹都表现出较高的趋同性。同时,在不同的生活环境及生理应激下,这些体色与斑纹又会出现显著的变化,给传统的形态分类带来巨大困扰,造成许多物种命名鉴定失误及分类混淆[1-5]。此外,石斑鱼亚科多个属之间的分类地位也存在争议或未明确。我国记载的石斑鱼亚科10个属中,部分资料将宽额鲈属(Promicrops)[2-3]、光腭鲈属(Anyperodon)[4,6]单独归为1属,但近期研究认为宽额鲈属、光腭鲈属的种类应归为石斑鱼属[1,7-10]。九棘鲈属(Cephalopholis)与侧牙鲈属(Variola)的分类地位也存在争议[1,6,11-12],下美鮨属(Hyporthodus)、烟鲈属(Aethaloperca)在石斑鱼亚科的分类地位也尚未明确。石斑鱼不乏多种海水经济养殖品种,物种分类不清,属间关系不明确不仅影响大众对石斑鱼物种的正确认知及资源利用,还会引起人工育种中出现近亲繁殖或错误配种问题,导致石斑鱼后代苗种质量的下降及种质资源的退化。
近年来,随着分子生物技术的发展,分子遗传标记在鱼类的分子鉴定及系统分类得到越来越广泛的应用,线粒体DNA不同基因在鱼类上已被验证可作为不同科、属、种分类鉴定的良好标记,解决了不少鱼类分类及系统演化问题[13-16]。16S rRNA基因进化速率较为保守,常用于种以上不同科、属阶元间的系统进化研究。细胞色素C氧化酶亚基I基因(COI)进化速率适中,可用于不同物种间的分类关系研究,其5´端一段长度为648 bp的片段,更是作为鉴定大多数鱼类物种的DNA条形码基因,能在分子水平上成功区分物种。将线粒体DNA标记应用于石斑鱼的系统分类研究,目前国内也有相关报道[9-10,17-18],但涉及的石斑鱼亚科种属均相对不多,同时缺乏关于COI基因的研究数据。国外Craig等[8]曾针对东太平洋及西大西洋分布的石斑鱼类进行较为系统的分子分类研究,但其缺乏印度西太平洋石斑鱼的种类。本研究基于16S rRNA、COI基因片段,对分布于西太平洋石斑鱼亚科10属40种鱼类进行系统进化树的构建,分析其不同种属间的系统分类关系,为进一步完善石斑鱼亚科鱼类分类研究提供分子依据。
2 材料与方法
2.1 样品采集与DNA的提取
本研究石斑鱼样品主要采集于广东、福建、海南等沿海省份的水产市场及渔港码头,最后共获得40种石斑鱼类,隶属于石斑鱼亚科10个属。采集标本根据《中国鱼类系统检索》[2]、《台湾鱼类志》[4]、《Grouper of the World》[1]等主要分类资料进行初步形态鉴定,样品取肌肉或鳍条,于95%酒精中固定,带回实验室-20℃冰箱保存,用于后期基因组DNA提取。采集样本信息见表1。
2.2 总基因组DNA的提取
取约50~80 mg的样品组织,利用动物组织DNA提取试剂盒(天根生化科技有限公司),按照其使用说明书进行提取。提取的总基因组DNA用100 μL灭菌水进行溶解,琼脂糖凝胶电泳检测DNA提取质量,-20℃保存备用。
2.3 PCR扩增和测序
本研究扩增的基因片段为16S rRNA及COI基因。参考相关文献,选择扩增16S rRNA基因的引物为 16S -F1:5´- CGC CTG TTT ATC AAA AAC AT -3´和16S -R1:5´- CCG GTC TGA ACT CAG ATC ACG -3´[19],扩增COI基因片段的引物为COI-F1: 5´- TCA ACY AAT CAY AAA GAT ATY GGC AC -3´和 COI-R1: 5´-ACT TCY GGG TGR CCR AAR AAT CA -3´[20]。PCR反应体系总体积为50 μL,其中包括PCR反应混合液 25 μL、无菌水 21 μL、上下游引物各1μL、DNA样品2 μL。PCR反应条件为94℃预变性5 min,94℃变性 30 s,55℃ 退火 30 s,72℃ 延伸1min,设置 35~38个循环,最后72℃再延伸5 min。PCR产物用1%琼脂糖凝胶电泳检测扩增结果,选取扩增成功的样品用PCR产物纯化试剂盒纯化回收后,送天一辉远生物技术有限公司进行测序。
2.4 数据分析方法
测序所得的序列先采用美国国家生物技术信息中 心 (National Center of Biotechnology Information,NCBI)的Blast工具进行相似性检索,验证序列为正确的COI基因序列。测序序列利用Clustal W[21]软件进行排序比对,核对修正,去掉首尾两端不规则冗余序列,得到一致序列进行后续分析。利用MEGA5.0[22]软件计算序列的碱基组成、转换颠换值、保守位点、变异位点、遗传距离等数据。系统进化树采用最大似然法,以鲈形目裸颊鲷科裸颊鲷属的星斑裸颊鲷及鲷科鲷属的黄锡鲷作为外类群进行构建,进化树采用重复抽样分析(Bootstrap Analysis)1 000次的方法检验各分支的置信度。
3 结果
3.1 基因序列分析
实验测得40种石斑鱼16S rRNA基因为580~610 bp,序列存在碱基插入与缺失,与作为外类群的黄锡鲷及星斑裸颊鲷的序列一起分析,经Clustal W排列比对,去掉冗余序列,得到566 bp一致序列,用MEGA 5.0软件进行分析。结果表明,序列中A、T、G、C碱基平均含量分别为29.2%、23.6%、22.8%、24.4%,其中A+T含量(52.8%)略高于G+C含量(47.2%)。除去外类群,保守位点394个(69.6%),变异位点172个(30.4%),简约性信息位点 140个(24.7%)。转换和颠换比值为3.269,转换大于颠换。
COI基因比对分析得到一致序列为651 bp,编码272个氨基酸。序列中A、T、G、C碱基平均含量分别为 24.1%、30.0%、18.3%、27.6%,其中 A+T含量(54.1%)略高于 G+C含量(45.9%)。如图 1所示,在651 bp COI基因编码序列中,密码子第1位4种碱基含量差异不大,相对最高的是G-1(31.1%);密码子第2位中,T-2含量(41.0%)最高,并明显高于其他3种碱基;密码子第3位中A-3含量最高(31.9%),G-3含量最低(8.3%),表现出明显的反G偏倚(图1)。在G+C含量中,密码子第1位点G+C含量(56.1%)高于第2和第3位(43.8%,37.8%)。此外,在长度为651 bp序列中,除去外类群,保守位点408个(62.7%),变异位点243个(37.3%),简约性信息位点227个(34.9%)。转换和颠换比值为3.111,转换大于颠换。

表1 试验材料的种类和来源Table1 Species and sources of experimental materials

图1 COI基因第1、第2、第3位密码子碱基组成Fig.1 Base composition of the 1st, 2nd and 3rd codon position of COI gene in Epinephelinae
3.2 种间与属间遗传距离
基于Kimura 2-Parameter模型利用MEGA5.0计算出各种之间16S rRNA以及COI的遗传距离(由于样品较多,表格数据较大,无法在同一页面显示,故这里不列出)。在16S rRNA基因中,除去外类群,40种石斑鱼类种内遗传距离为0~0.003,平均遗传距离为0.001;种间遗传距离范围为0.002~0.168,平均遗传距离为0.090,其中豹纹鳃棘鲈与蠕线鳃棘鲈遗传距离最小(0.002),其次是白背石斑鱼与花点石斑鱼,云纹石斑鱼与褐石斑鱼,均为0.007;横斑鳃棘鲈与青星九棘鲈、六斑九棘鲈遗传距离最大,均为0.168,其次是横斑鳃棘鲈与马拉巴石斑鱼,为0.167;在石斑鱼属内种间遗传距离中,21种石斑鱼属鱼类遗传距离范围是0.007~0.067,最小的是白背石斑鱼与花点石斑鱼(0.007),其次是赤点石斑鱼与青石斑鱼,以及细点石斑鱼与蓝棕石斑鱼(0.011);最大是吊桥石斑鱼与玳瑁石斑鱼(0.067),其次是花点石斑鱼与吊桥石斑鱼(0.063)。在6种鳃棘鲈中,豹纹鳃棘鲈与蠕线鳃棘鲈最小(0.002),蓝点鳃棘鲈与点线鳃棘鲈,以及蓝点鳃棘鲈与横斑鳃棘鲈最大(0.050),5种九棘鲈中青星九棘鲈与六斑九棘鲈最小(0.015),横带九棘鲈与青星九棘鲈最大(0.086)。
在COI基因中,40种石斑鱼种内遗传距离为0~0.006,平均遗传距离为0.003;种间遗传距离范围为0.016~0.245,平均遗传距离为0.172,可知COI基因序列差异性比16S rRNA要大,碱基变异数目更多。其中遗传距离最大的是宽额鲈与侧牙鲈,遗传距离最小的是云纹石斑鱼与褐石斑鱼;在石斑鱼属内部,21种石斑鱼属鱼类遗传距离范围为0.030~0.192,最小的是云纹石斑鱼与褐石斑鱼(0.030),其次是斜带石斑鱼与马拉巴石斑鱼(0.036);最大是玳瑁石斑鱼与纵带石斑鱼(0.192),其次是玳瑁石斑鱼与蓝棕石斑鱼(0.181)。在6种鳃棘鲈中,豹纹鳃棘鲈与蠕线鳃棘鲈最小(0.016),点线鳃棘鲈与斑鳃棘鲈最大(0.123),5种九棘鲈中青星九棘鲈与六斑九棘鲈最小(0.043),横带九棘鲈与青星九棘鲈最大(0.180)。
同时,我们汇总了石斑鱼亚科10个属属间的遗传距离范围(表2,表3)。在16S rRNA中,遗传距离范围最大是鳃棘鲈属与九棘鲈属(0.147~0.168),而石斑鱼属与鸢鮨属遗传距离范围最小(0.036~0.054);而在COI基因中,遗传距离范围最大是侧牙鲈属与驼背鲈属(0.216~0.238),而遗传距离范围最小是石斑鱼属与驼背鲈属(0.101~0.176)。另外,基于属间遗传距离比较发现,部分属与石斑鱼属的遗传距离都比较小,甚至小于石斑鱼属属内种间的部分遗传距离(石斑鱼内部属内种间遗传距离分别为16S rRNA:0.007~0.168,COI:0.030~0.192),如驼背鲈属(16S rRNA:0.044~0.071,COI:0.101~0.176)、下美鮨属(16S rRNA:0.036~0.061,COI:0.108~0.170)、光腭鲈属(16S rRNA:0.037~0.067,COI:0.104~0.178)、鸢鮨属(16S rRNA:0.036~0.054,COI:0.142~0.203)、宽额鲈属(16S rRNA:0.039~0.075,COI:0.123~0.195)等与石斑鱼属的遗传距离都相对较小,显示出这4个属与石斑鱼属可能有比较接近的亲缘关系。

表2 基于Kimura-2模型石斑鱼亚科鱼类各属间16S rRNA序列遗传距离Table2 Pairwise distances among genera of Epinephelinae for 16S rRNA sequences based on Kimura-2 model
3.3 石斑鱼亚科鱼类分类及分子系统树
基于16S rRNA,COI以及16S rRNA+COI整合序列,以黄锡鲷及星斑裸颊鲷为外类群,利用最大似然法构建系统进化树。如图2至图4所示,3棵进化树拓扑结构差异不大,其中16S rRNA+COI整合序列与单个COI基因构建的系统树结构基本一致,16S rRNA基因构建的树在部分物种间的聚类关系与前两者有少量差异。进化树上,同种石斑鱼不同个体均能紧密聚在一起,形成单系分支。鉴于个体样品数量多,进化树上无法一一显示,故进化树仅展示40种石斑鱼亚科各物种聚类关系。这里主要分析16S rRNA+COI整合序列构建的系统进化树。除去外类群,石斑鱼亚科10个属大致形成4个分支,其中第1个巨大分支由21种石斑鱼属鱼类与驼背鲈属、光腭鲈属、宽额鲈属、下美鮨属及鸢鮨属6个属组成,位于进化树的顶部,石斑鱼属鱼类并未形成单系分支。该分支内部又形成两个类群,类群I包括莹点石斑鱼、珊瑚石斑鱼等10种石斑鱼属与驼背鲈属、光腭鲈属、宽额鲈属、下美鮨属、鸢鮨属的种类组成;类群II主要包括玳瑁石斑鱼、三斑石斑鱼等11种石斑鱼属鱼类;第2分支由九棘鲈属与烟鲈属组成;第3分支由侧牙鲈属两个种类组成;最后一支为鳃棘鲈属的6个种类形成的单系,位于进化树的基部。
4 讨论
4.1 石斑鱼亚科序列特征及遗传距离分析
根据16S rRNA及COI基因序列特征分析,本研究中石斑鱼亚科鱼类16S rRNA及COI基因碱基特征与其他鱼类基本相似,变化不大[13,23-24],两基因平均转换和颠换比值均大于2,序列碱基替代还没有达到饱和,可用于后期系统发育分析。遗传距离中,基于COI基因石斑鱼亚科鱼类种内遗传距离(0~0.006)均小于最小种间遗传距离(0.016),而在16S rRNA基因中,蠕线鳃棘鲈与豹纹鳃棘鲈两物种间遗传距离(0.002)却小于个别物种的种内遗传距离(0~0.003),显示蠕线鳃棘鲈样品与豹纹鳃棘鲈16S rRNA基因水平十分接近。本研究中蠕线鳃棘鲈共采集到两个个体,豹纹鳃棘鲈4个个体,但两种鳃棘鲈种内遗传距离均为0,根据形态特征鉴别,蠕线鳃棘鲈与豹纹鳃棘鲈体型相似,体色也均为红色,但两种鱼体表斑点具明显差别,蠕线鳃棘鲈体表分布暗色圆形斑点以及短条状的斑纹,而豹纹鳃棘鲈密布闪亮的浅蓝色斑点,据此特征可区分两者[1-2,4,25]。至于为何两者16S rRNA序列如此接近,有可能是两种石斑鱼在自然界存在杂交导致基因渗透,也可能是16S rRNA序列本身在进化上比较保守,近缘物种之间差异非常小,序列相似性很高所致[26]。本研究采集的蠕线鳃棘鲈与豹纹鳃棘鲈样品,基于两者形态差异显著,16S rRNA种内遗传距离为0,COI基因种间遗传距离也远大于其种内遗传距离,故本研究将蠕线鳃棘鲈与豹纹鳃棘鲈暂以两独立物种对待。
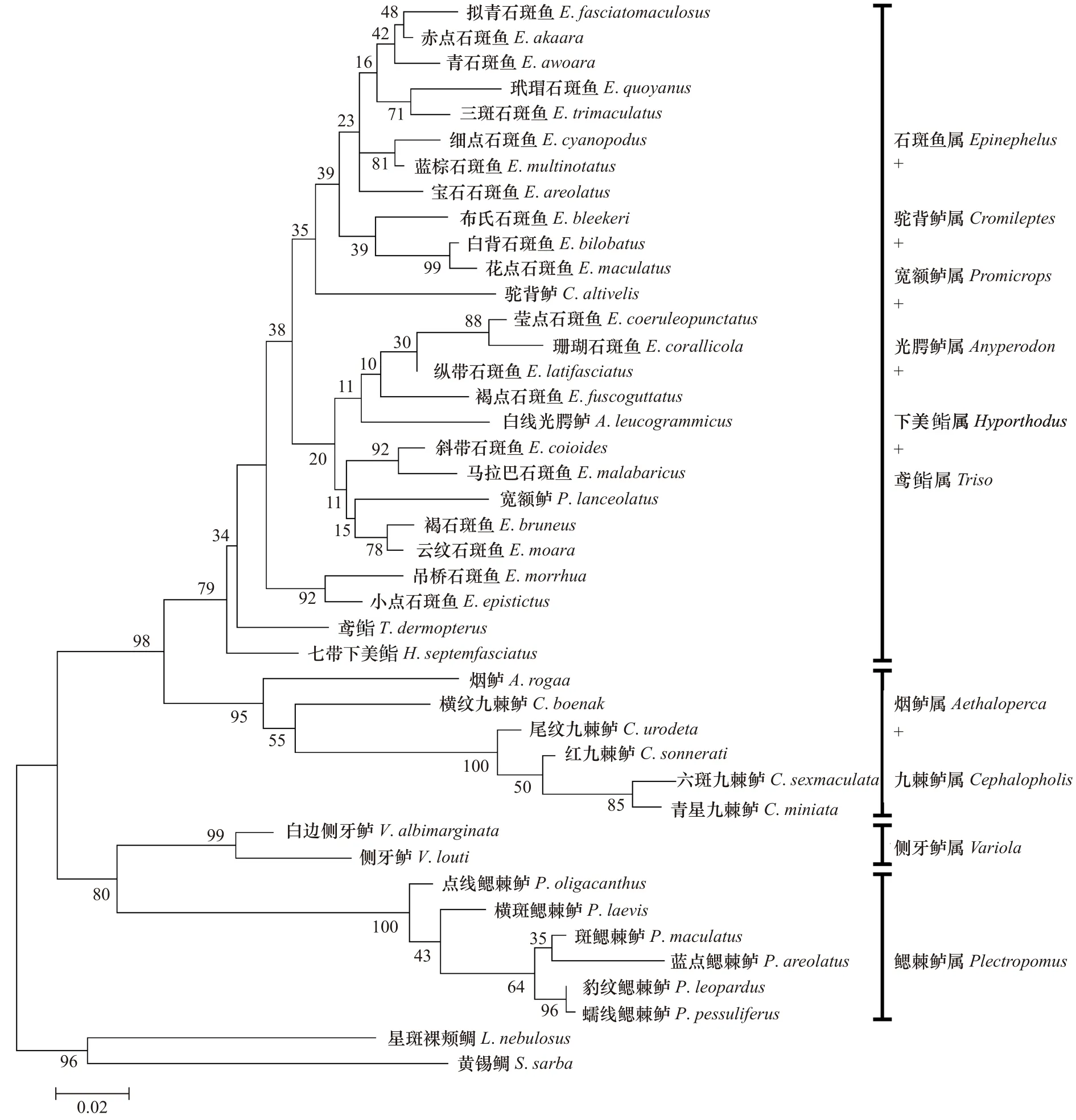
图2 40种石斑鱼亚科鱼类基于16S rRNA序列利用最大似然法构建的分子系统进化树Fig.2 Molecular phylogenetic trees of 40 Epinephelinae fish based on 16S rRNA sequences constructed by maximum likelihood method
4.2 鳃棘鲈属的系统分类关系
构建的系统进化树上,鳃棘鲈属最先分化,进化地位最原始,这与前人不管是形态学还是分子系统学研究的结果一致。形态上,鳃棘鲈属背鳍鳍棘6~8枚,石斑鱼亚科其他属背鳍鳍棘9~11枚,是石斑鱼亚科中鳍棘数最少的1个属[2-4]。Leis[12]以鳃棘鲈属幼体的腹鳍鳍棘结构,眶上和前鳃盖骨棘,鳍条分支和色素变化4大特征,揭示鳃棘鲈属是石斑鱼亚科的原始类群。Craig等[8]首次基于16S rRNA对南北美洲沿岸的42种石斑鱼类系统进化研究,从分子水平揭示鳃棘鲈比其他石斑鱼地位原始。丁少雄等[9]、庄轩等[10]研究团队分别采用16S rRNA、Cyt b基因对我国近30种石斑鱼的分类研究,也认为鳃棘鲈属的分化时间早于其他石斑鱼类。本研究基于16S rRNA、COI分析同样得出上述结果。鳃棘鲈属全球共7种,6种分布于西太平洋,1种分布于印度洋[25]。本研究把西太平洋分布的6种鳃棘鲈属全部采集,系统进化树上,6种鳃棘鲈属聚成一个单系,显示鳃棘鲈属为一单系群。

图3 40种石斑鱼亚科鱼类基于COI序列利用最大似然法构建的分子系统进化树Fig.3 Molecular phylogenetic trees of 40 Epinephelinae fish based on COI sequences constructed by maximum likelihood method
4.3 九棘鲈属、烟鲈属与侧牙鲈属的系统分类关系
传统形态学认为九棘鲈属应该作为一个亚属归为石斑鱼属,但近代不少研究认为九棘鲈属不管在形态上(背鳍鳍棘数、背鳍支鳍骨数目)[6]、个体发育(黑色素细胞数目与位置变动)[12]还是分子水平上[8-10,27-28],均应该是一个独立的属。本研究系统进化树上,九棘鲈属种类单独聚为一支,并独立于石斑鱼属类群之外,支持九棘鲈属为独立一属的观点。

图4 40种石斑鱼亚科鱼类基于16S rRNA与COI整合序列利用最大似然法构建的分子系统进化树Fig.4 Molecular phylogenetic trees of 40 Epinephelinae fish based on combined 16S rRNA and COI sequences constructed by maximum likelihood method
侧牙鲈属的分类地位介于鳃棘鲈属与九棘鲈属之间,与前人的研究结果一致[8-10,26,28],侧牙鲈属的分化晚于鳃棘鲈属,早于九棘鲈属。基于16S rRNA及COI基因序列分析,在16S rRNA中,侧牙鲈属与九棘鲈属(0.126~0.142)的遗传距离比鳃棘鲈属要大(0.108~0.131),而在 COI基因(0.191~0.219)中却比鳃棘鲈属要小(0.205~0.237)。形态上,侧牙鲈属与九棘鲈属背鳍均具有9个鳍棘,而鳃棘鲈为6~8枚鳍棘,庄轩等[10]基于Cyt b序列分析也得出侧牙鲈属与九棘鲈属遗传距离较鳃棘鲈要小。故综合分析,本研究也倾向于认为,相对于鳃棘鲈属,侧牙鲈属与九棘鲈属关系更接近。
烟鲈属在分类学上属于单型属[4],仅烟鲈属一种。烟鲈属全身棕褐色,有时腹部具一道白斑,根据其前鳃盖骨下缘无向前棘,胸鳍第5或第6软鳍条最长等特征与其他属的种类区分开[4]。进化树上,烟鲈属与九棘鲈属紧密聚为一支,显示两者关系十分接近。Craig和Hastings[27]构建的石斑鱼亚科系统进化树也揭示烟鲈属聚类于九棘鲈属中,Zhuang等[29]基于线粒体基因组全长分析22种石斑鱼也表明烟鲈属与九棘鲈属关系最近。形态上,烟鲈属与九棘鲈属同样具有9枚背鳍鳍棘的主要性状。但由于九棘鲈属样品数量有限,在聚类关系上,烟鲈属与九棘鲈属是两个平行进化的分支,还是烟鲈属位于九棘鲈属内部,本研究还不能给予确切结果,需要后期采集更多九棘鲈属种类样品进一步分析才能明确。
4.4 宽额鲈属、光腭鲈属与下美鮨属的系统分类关系
宽额鲈在我国传统的一些分类资料上是独立为一个属[2-3],Smith[7]根据骨骼发育特征将宽额鲈属作为一个亚属归入石斑鱼属,而近期国内外关于宽额鲈属的分子系统学研究认为宽额鲈属与石斑鱼属有很近的亲缘关系,Craig等[8]、丁少雄等[9]、庄轩等[10]等前人关于石斑鱼亚科分子系统分类研究,宽额鲈聚类于石斑鱼属内部,建议宽额鲈归入石斑鱼属。本研究构建的3棵系统进化树,宽额鲈均位于石斑鱼属分支内部,与斜带石斑鱼、马拉巴石斑鱼、云纹石斑鱼、褐石斑鱼关系最近,同意前人认为宽额鲈归入石斑鱼属的观点。
白线光腭鲈腭骨无齿、两颌缝合部两侧无膨大犬齿,可与石斑鱼亚科其他属区分,独立为单型属[4,6],本研究构建的3棵进化树,白线光腭鲈均位于石斑鱼属内部,没有形成单一分支,独立于石斑鱼属之外。Zhuang等[29]基于线粒体全长序列分析22种石斑鱼类系统分类关系也揭示白线光腭鲈聚类于石斑鱼属内部。陈艺燕等[17]基于Cyt b部分序列研究10种石斑鱼类系统发育关系,白线光腭鲈与石斑鱼紧密聚在一起。形态上,白线光腭鲈与石斑鱼属比较,两者也具有部分相似的特征,如背鳍鳍棘为11条,背鳍支鳍骨只有两节等等[4]。本结果倾向于支持白线光腭鲈归入石斑鱼属的观点。
下美鮨属的七带下美鮨在传统分类学中又叫七带石斑鱼(Epinephelus septemfasciatus),归为石斑鱼属[2-3]。但形态上它跟石斑鱼属也存在一定的区别:七带下美鮨腹鳍一般在胸鳍的正下方或者前方,石斑鱼属腹鳍一般在胸鳍后方。而近期的分子系统学研究中,下美鮨属与石斑鱼属的进化关系非常接近[27-28],认为其应该归为石斑鱼属。在本研究进化树同样揭示七带下美鮨与石斑属鱼类有很近亲缘关系,七带下美鮨紧密聚类于石斑鱼属内部,基因序列间的差异也比较小(16S rRNA:0.036~0.061;COI:0.108~0.170),并小于大部分石斑鱼属内种间的遗传差异(16S rRNA:0.007~0.168;COI:0.030~0.192),该结果支持近期认为七带下美鮨归为石斑鱼属的观点。
4.5 驼背鲈属、鸢鮨属的系统分类关系
驼背鲈头部背缘强烈凹入,驼背状隆起,后鼻孔垂直裂孔状,背鳍鳍棘为10等独特外观,明显区别于石斑鱼亚科其他种类,目前分类资料也将其单独归为一属—驼背鲈属[2-4]。本研究中,驼背鲈属与石斑属关系十分接近,进化树上聚类于石斑鱼属内部,遗传距离差异也比较小(16S rRNA:0.044~0.071;COI:0.101~0.176),并小于部分石斑鱼属内种间的遗传差异,显示其与石斑属亲缘关系密切,该结果亦与前人分子系统研究结果一致[8-10,29]。但形态上,驼背鲈与石斑鱼属鱼类差异十分显著,这种外部形态与分子水平的差异可能是分子进化速率与表型进化速率不一致,表型进化出现趋异现象所致[30]。丁少雄等[9]认为其可能是石斑鱼属内的一个特化类群,或是某一共同祖先平行演化的结果,但还需更多的形态学及分子系统学的数据进一步的分析才能确定。
鸢鮨属也是单型属,仅鸢鮨一种,体型呈椭圆形,棕色,两眼间隔区域显然隆突,与石斑鱼属鱼类相比,具有较多的背鳍及臀鳍鳍条[4]。但本研究所有进化树均揭示鸢鮨位于石斑鱼属内部,与石斑鱼属关系密切。Schoelinck等[28]、Zhuang等[29]对石斑鱼系统发育研究分析也认为,鸢鮨与石斑属鱼类关系非常接近。但基于形态的明显差异,鸢鮨属应归入石斑鱼属,还是与驼背鲈一样可能是特化类群,还需后续更深一步的研究。
5 结论
综上所述,本研究基于16S rRNA、COI所分析的石斑鱼亚科10属40种鱼类分子系统进化关系中,鳃棘鲈属分类地位最原始,位于进化树基部;侧牙鲈属的进化地位介于鳃棘鲈属与九棘鲈属之间;烟鲈属与九棘鲈属关系较近,两者聚为一支;石斑鱼属的进化地位最高,位于进化树顶部,形成两个平行分支,但是石斑鱼属种类未能聚成一个单系。驼背鲈属、鸢鮨属、下美鮨属、光腭鲈属及宽额鲈属与石斑鱼属种类聚一起,显示其与石斑鱼属有很近的亲缘关系,部分可能是石斑鱼属的特化类群。研究结果可为石斑鱼亚科鱼类分类提供分子水平依据,也为后续相关鱼类分子系统进化研究提供参考。
