贵州麻江分布小弧斑姬蛙的分子鉴定
2020-07-20韩丹余文华张蓉
韩丹 余文华 张蓉

摘要:姬蛙是一大类常见的农田物种,通常个体较小,蝌蚪间差异不明显,难以准确鉴定。本研究对采自贵州省黔东南州麻江县姬蛙类蝌蚪标本进行DNA提取,使用PCR技术扩增线粒体16S rRNA基因片段,参照GenBank中同源序列的系统发育关系对其进行初步分子鉴定。结果表明,标本16S rRNA基因片段与27条小弧斑姬蛙(Microhyla heymonsi)同源序列聚为一个单系,该单系内部序列间遗传距离小于本研究所用参考序列的种间遗传距离,因此,初步鉴定标本为小弧斑姬蛙蝌蚪。同时,本研究中小弧斑姬蛙支系的序列间遗传距离略高于所有参考序列的种内遗传距离,并且在單倍型网络中被分割为6个独立网络,暗示分布广泛的小弧斑姬蛙种内存在较大的支系分化。另外,本研究对参考序列中的可疑序列进行了初步探讨。
关键词:小弧斑姬蛙;16S rRNA基因;克隆;分子鉴定;可疑序列
小弧斑姬蛙(Microhyla heymonsi),隶属姬蛙科(Microhylidae)、姬蛙属(Microhyla)[1],是常见的农田蛙类。因其具有多样的环境适应能力,而广泛分布于中国、印度、缅甸、泰国、老挝、越南、柬埔寨、马来半岛、印度尼西亚等地区[1-2]。国内主要分布于四川、重庆、云南、贵州、河南、安徽、江苏、浙江、江西、湖南、福建、台湾、广东、海南、广西等地区[1]。小弧斑姬蛙在贵州省内多地均有分布,如绥阳、仁怀、赤水、大方、金沙、江口、印江、松桃、贵阳、贵定、荔渡、罗甸、雷山、兴义、安龙、德江、剑河等地区[3]。然而,目前并没有明确的文献记载该物种在麻江县是否有分布。2016年6月,笔者于贵州省黔东南州麻江县坝芒乡采集到疑似小弧斑姬蛙蝌蚪标本3号,但因蝌蚪较小,形态特征不明显,且与贵州分布的其他姬蛙蝌蚪极为相似,因此有必要对该标本使用分子鉴定手段作进一步的物种鉴定。本研究拟对所采集的姬蛙蝌蚪标本进行DNA提取,扩增其线粒体16S rRNA基因(以下简称16S基因),并进行系统发育分析从而对其物种归属进行初步鉴定。
1 材料与方法
1.1 标本信息
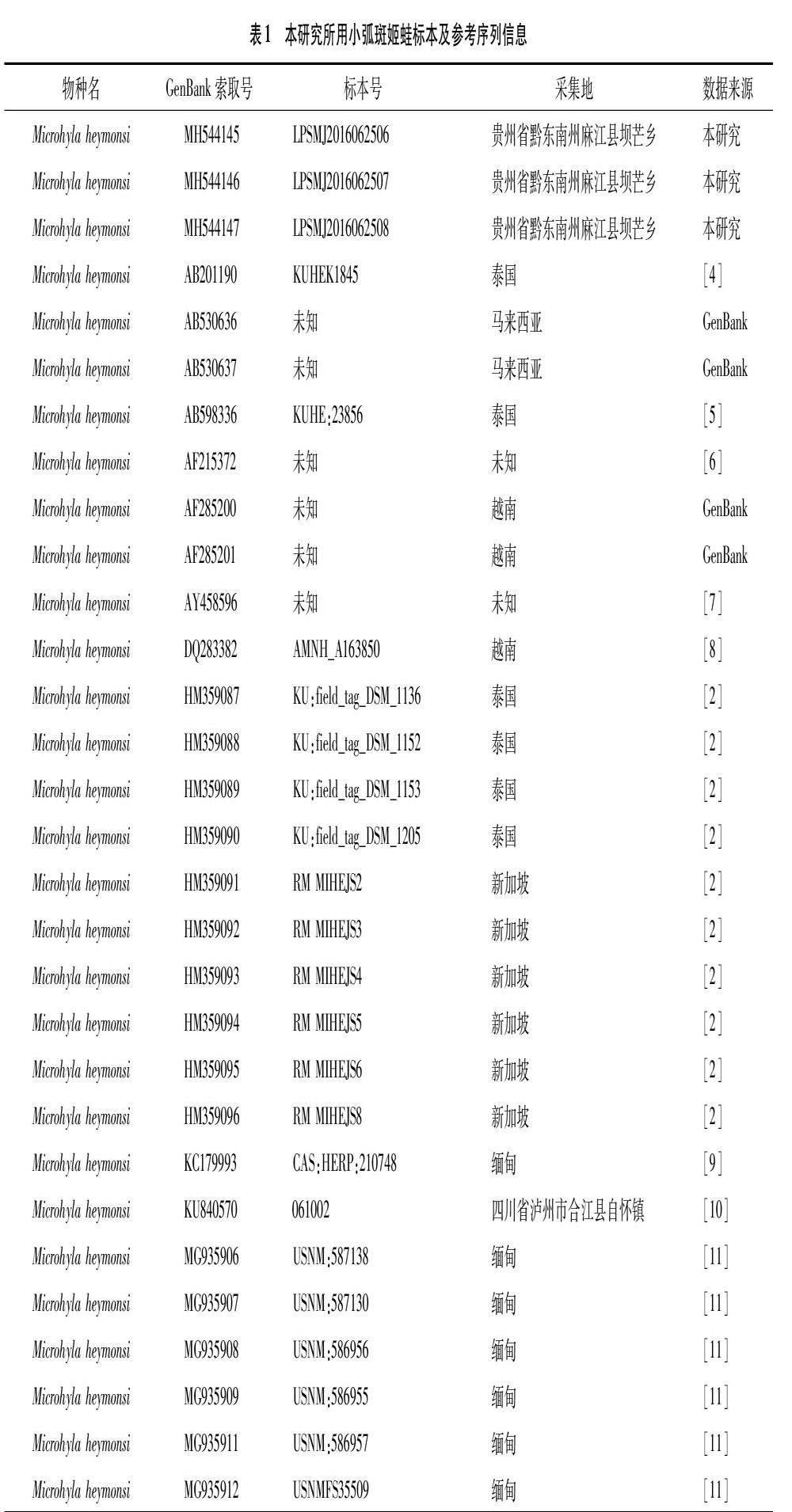
小弧斑姬蛙标本为2016年6月25日采自贵州省黔东南州麻江县坝芒乡的3个蝌蚪幼体,保存于六盘水师范学院生物科学与技术学院动物标本馆。参考序列[4-11]见表1。
1.2 总DNA提取及线粒体16S rRNA基因片段扩增
取蝌蚪尾部肌肉,使用动物组织DNA提取试剂盒(Foregene,DE-05011:250 Preps)提取总DNA。扩增引物为1对脊椎动物线粒体16S基因片段扩增通用引物(P7/P8)[12-13]。PCR反应条件与华西雨蛙、寒露林蛙同源基因扩增条件相同[13-14],PCR产物经1%琼脂糖凝胶电泳检测后送至成都擎科梓熙生物技术有限公司测序。
1.3 参考基因序列下载及系统发育分析
将所测得的小弧斑姬蛙线粒体16S基因序列经过上传GenBank进行搜索比对(Mega Blast)获得100条参考序列,与本研究所得基因序列一起构成数据集A。用Muscle软件[15]对序列进行比对,在Mtga X[16]中辅以人工校对并将首尾不整齐片段删除。进行系统发育分析时不预先设定外群,构建无根系统发育树。使用jModel Test[17]中赤池信息量准则(akaike information criterion,简称AIC)筛选适合数据集A的最优模型,并使用相应的最优模型树(best model tree)作为拓扑结构基准。根据AIC,适合该数据集的模型为GTR+I+G模型,其中 G=0.885,I=0.605。使用raxmlGUI软件[18]构建最大似然树(maximum likelihood tree,简称ML tree),自举检验支持率>70%表明该支系关系得到充分解决。使用Mrbayes 3.2[19]构建贝叶斯树(bayesian inference tree,简称BI tree),碱基替换模型设置为GTR+G+I,以后验概率(posterior probability,简称PP)来表示各分支的可信度(支持率),贝叶斯后验概率≥95%说明其支系支持率得到充分解决。使用Mega X软件构建邻接树(neighbor-joining tree,简称NJ tree)和最大简约树(maximum parsimony tree,简称MP tree),自举检验支持率>70%表明该支系关系得到充分解决,支持率在50%~70%之间为中度支持,否则视为未解决。
1.4 遗传距离的计算
选用软件Mtga X,分析数据集A中各序列间差异,依据Kimura 2-parameter模型计算两两序列间的遗传距离以及主要支系间的平均遗传距离。使用R语言基础包(base)绘制种间及种内遗传距离数据的密度分布曲线,探明小弧斑姬蛙及近缘物种在16S基因上的种间及种内遗传距离的主要分布区域。
1.5 单倍型网络构建
根据双重单系法[5,20],在数据集构建的系统发育树上,以自测序列为起点向根部回溯2个单系中间节点,从而得到包含自测序列及其近缘物种同源基因序列构成数据集B。使用R语言程序包haplotypes[21]分析数据集B的单倍型类型并绘制单倍型网络(图1-A)。设置简约上限(parsimony limit)时,默认值为0.95,考虑到要将不同的物种划分到独立的网络中去,本研究中简约上限设置为0.97。
2 结果与分析
2.1 PCR扩增及测序结果
经1%琼脂糖凝胶电泳检测,引物P7/P8扩增到550 bp左右的基因片段。经测序,获得长度为544~555 bp DNA序列(GenBank索取号为MH544145~MH544147)。序列碱基组成存在明显的偏向性:AT含量较高(A含量为31.1%,T含量为25.5%);GC含量较低(G含量为20.9%,C含量为22.6%)。所扩增最长序列MH544146(555 bp),对应小弧斑姬蛙线粒体基因组的(GenBank索取号为AY458596)2 161~2 703 bp区间位置,对应小弧斑姬蛙16S rRNA全基因(位于线粒体基因组AY458596的1 288~2 861 bp区域)874~1 416 bp区间位置,因此扩增目的序列为小弧斑姬蛙16S rRNA编码基因的下游区域。
2.2 序列及其系统发育分析结果
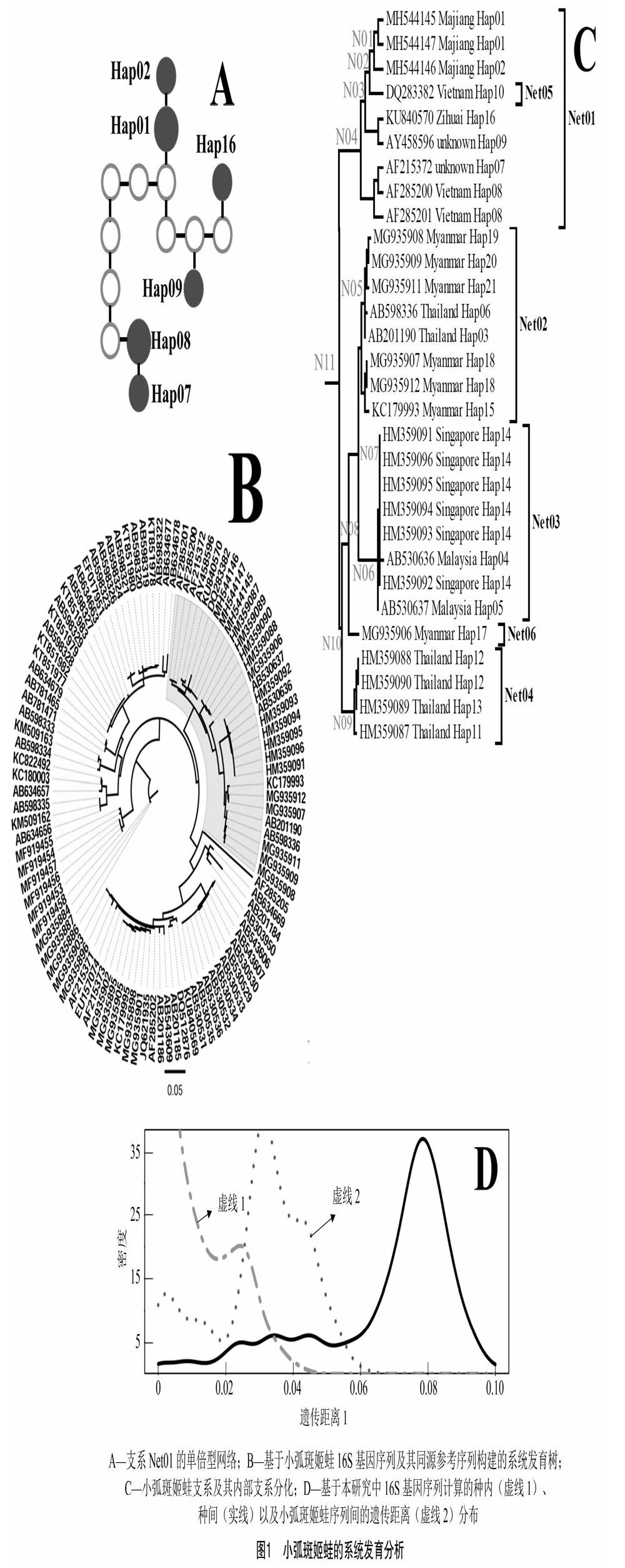
将所测得16S基因序列与100条同源序列一起构建的系统发育树中(图1-B),3条自测序列与27条小弧斑姬蛙同源序列聚为1个单系(小弧斑姬蛙支系,图1-C中节点N11),同时得到NJ(97%)、MP(90%)、ML(88%)以及BI(100%)等方法的较高支持率支持(表2)。在其内部,进一步分化为5个主要支系(节点N04、N05、N06、N09所在支系以及单序列支系MG935906)。N05、N06、N09均得到了4种系统发育推断方法较高支持(表2)。本研究中3条自測序列与自怀、越南等种群的6条序列聚为1个支系(N04),但支持率较低(表2)。
2.3 单倍型网络
提取小弧斑姬蛙支系序列构建数据集B,以之构建单倍型网络,在将简约上限设置为默认值0.95时,并未将所有序列包括在同一个单倍型网络中,其中支系N06中的3条序列(AB530636、AB530637、HM359091)单独构成1个独立网络。为进一步将主要支系都分割成独立的网络,更清晰地观察自测序列单倍型所属网络情况,根据系统发育树的主要支系分化的拓扑结构,将简约上限设置为 0.97,以得到更多的相互独立的单倍型网络(6个单倍型网络分别为Net01、Net02、Net03、Net04、Net05,Net06,图1-C)。然而,当所有支系都独立成单独的单倍型网络时,支系N01绝大部分序列构成单倍型网络Net01(图1-A),而序列DQ283382单独构成一个单倍型网络Net05(图1-C)。初步推定该序列为一条可疑序列,为探究其对系统发育关系构建的影响,从数据集A中剔除DQ283382,重新构建序列间的系统发育关系,探究该序列对系统发育树拓扑结构和节点支持率的影响(表2)。
2.4 遗传距离
基于16S基因序列的遗传距离表明,小弧斑姬蛙支系内遗传距离为0.0%~1.6%,其内部支系Net01、Net02、Net03、Net04的内部序列间平均遗传距离分别为1.12%、0.87%、0.15%、0.33%,亚支系间的遗传距离为3.6%(2.2%~5.6%);亚支系和非小弧斑姬蛙序列间的遗传距离为7.7%(7.2%~8.7%)。种间及种内遗传距离数据的密度分布曲线(图1-D)表明,相比所有序列种间遗传距离分布曲线,小弧斑姬蛙支系内部序列间的遗传距离分布曲线和种内遗传距离分布曲线重合较多,但总体高于种内遗传距离。
3 讨论
3.1 小弧斑姬蛙的分子鉴定
本研究系统发育分析结果表明,3条自测序列与27条小弧斑姬蛙同源序列聚为1个单系,既使去掉可疑序列(DQ283382)后,仍与26条小弧斑姬蛙同源序列聚为1个单系(表2)。3条自测序列与来自我国自怀以及越南的小弧斑蛙同源序列聚为1个亚支系,内部序列间平均遗传距离分别为1.12%,远低于本研究中所有序列的种间遗传距离(图1-D)。因此,初步判定本研究所采集蝌蚪标本属于小弧斑姬蛙(Microhyla heymonsi)。但本研究涉及的麻江、自怀种群均距离模式产地台湾[3]较远,且缺乏其他种群数据(台湾种群、东部种群等),因此鉴定结果有待进一步证实。
3.2 小弧斑姬蛙内部支系分化
在本研究中,小弧斑姬蛙支系进一步分化为6个亚支系,各支系在单倍型网络分析中被分割为独立的网络(排除1条可疑序列后,仍有5个支系)。且部分支系单系性在多种系统发育分析方法中得到较高的支持率,如支系Net02、Net03。亚支系间的遗传距离与种内距离不完全重合,偏高,但并没有超越种间距离(图1-D),说明小弧斑姬蛙物种内部种群间有了较大的遗传分化。考虑其较为广阔的分布范围[3],这种分化可能在一定程度上是由于种群间相距较远,基因交流频率较低所致,详细的种内支系分化关系以及分化水平有待更为充分的抽样研究支撑。
3.3 可疑序列剔除方法初探
在本研究使用的参考序列中DQ283382在系统发育关系重建中聚在支系N04内部,但在单倍型网络构建中,其并未与该支系的其他序列链接为一个共同的网络(Net01),而是独立成为1个单独网络(Net05)。鉴于该序列所在种群未有其他同源序列[4](生物重复),初步判定为一条可疑序列。为探究该可疑序列对于系统发育关系重建的影响,本研究将其剔除后重新构建的系统发育树与未剔除时的系统发育树作对比。结果(表2)表明,剔除该序列后,不同方法构建的系统发育树的拓扑结构更趋一致,如在剔除前,节点N03同时得到了MP、ML、BI方法的支持,但没有得到NJ的支持,在剔除后,同时得到了4种方法的支持;又如节点N10的聚类关系只得到了ML、BI的支持,剔除DQ283382后,增加了MP方法的支持。另外,剔除序列DQ283382后,系统发育树的大多数节点支持率都有所提高。例如N01节点,MP的支持率由49%上升为94%,BI的支持率由<50%上升至82%;又如N03节点,ML树的支持率由60%上升到75%;N04节点的NJ树的支持率由57%上升至73%。
综上所述,可疑序列DQ283382在一定程度上影响了系统发育关系的重建,而对比单倍型网络与系统发育树拓扑结构的一致性,提供了识别类似可疑序列的一个视角,当然其适用性还有待更多的案例研究加以检验。
参考文献:
[1]费 梁,叶昌媛,江建平. 中国两栖动物及其分布彩色图鉴[M]. 成都:四川科学技术出版社,2012:568-569.
[2]Sheridan J A,Bickford D,Feng K,et al. An examination of call and genetic variation in three wide-ranging Southeast Asian anuran species[J]. The Raffles Bulletin of Zoology,2010,58(2):369-379.
