不同施钾量对淀粉型甘薯光合特性、淀粉含量及产量的影响
2020-07-20密扬陈剑秋王瑞霞
密扬 陈剑秋 王瑞霞

摘要:为明确不同施钾量对淀粉型甘薯淀粉含量的调控效应及其与光合特性的关系,选取徐薯18和商薯19这2个淀粉型甘薯代表品种布设2年田间试验,设CK(不施肥)、K0(不施钾肥)、K75(施K2O 75 kg/hm2)、K150(施K2O 150 kg/hm2)和K225(施K2O 225 kg/hm2)5个处理,研究施钾量对甘薯淀粉产量和淀粉含量等的影响,利用逐步回归分析等方法探究淀粉合成与光合指标的相互关系。结果表明,徐薯18的3个施钾处理淀粉产量分别提高了16.5%、25.7% 和28.8%;商薯19的3个施钾处理淀粉产量分别提高了5.9%、11.9%和34.9%。2个甘薯品种的总淀粉含量随着施钾量的升高而呈显著升高趋势(P<0.05),其中徐薯18各施钾处理分别较K0处理提高0.4、1.2、2.4百分点,商薯19各施钾处理分别较K0处理提高0.4、0.8、2.1百分点。增加钾肥施用量能显著提高甘薯直链淀粉含量,进而提高甘薯直链淀粉含量/支链淀粉含量。随着施钾量的增加,甘薯的SPAD值、净光合速率(Pn)、最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率(ΦPSⅡ)、光化学淬灭系数(qP)和电子传递速率(ETR)整体上呈上升趋势,甘薯淀粉含量与SPAD值、Pn、Fv/Fm、qp、ETR呈显著正相关,与钾积累量呈极显著正相关(r=0.96,P<0.01)。综合甘薯产量和淀粉含量因素,推荐徐薯18的钾肥用量为150 kg/hm2,商薯19的钾肥用量为225 kg/hm2。研究结果可为淀粉型甘薯的钾肥高效利用提供理论基础。
关键词:淀粉型甘薯;产量;钾肥用量;钾肥利用;光合特性
甘薯[Ipomoea batatas(L.)Lam.]作为一种重要的粮食作物和工业原料,在农业和相关产业的发展中具有重要意义[1]。淀粉是甘薯块根的重要组分,其积累量直接决定其是否高产[2-4]。同时,淀粉含量、直链和支链淀粉的含量及比例、淀粉粒粒径大小及分布特征等性状与块根淀粉性能密切相关[5-6]。增施钾肥不仅能促进甘薯体内糖分的运输[7],提高干物质在块根中的分配率,增加干物质生产量和块根产量,还能促进淀粉合成,进而增加淀粉粒数[8]。其他研究发现,施用钾肥能提高甘薯关键生育期蔗糖合酶(SS)、颗粒结合淀粉合成酶(GBSS)和腺苷二磷酸葡萄糖焦磷酸化酶(ADPGase)等酶的活力,增加甘薯块根淀粉含量[9-10]。以往研究多通过酶学角度揭示施钾对甘薯淀粉含量的调控效应,而施钾条件下光合特性与甘薯淀粉含量的相互作用却鲜有报道。
甘薯是绿叶植物,可通过光合作用补充新陈代谢所需的能量。研究表明,叶片的光合指标、荧光指标和叶绿素含量等可作为衡量甘薯植株光合能量强弱的参考依据,而钾素往往是影响植株光合能力的重要因素[11-13]。因此,研究不同鉀肥用量对甘薯叶片光合指标和叶绿素荧光参数等的影响,对于揭示施钾条件下光合特性与甘薯淀粉含量的相互关系有重要的意义。本研究立足于田间试验,选取高淀粉型和中淀粉型2个甘薯品种,结合淀粉型甘薯生长特性,探究不同施钾量对甘薯淀粉含量、钾素吸收积累量和光合特性的影响,探讨施钾提高甘薯淀粉含量的光合机制,以期为甘薯产业的钾肥用量选择提供理论指导。
1 材料与方法
1.1 试验地概况
选用2种淀粉型甘薯代表品种(徐薯18和商薯19)于2017年5月20日在山东省临沂市沂水县向阳村进行连续2年田间试验。布设试验前,各小区分别以5点法采集0~20 cm深土壤样品,风干后采用 1.0 mol/L NaOH扩散法测碱解氮含量,采用 0.5 mol/L NaHCO3浸提-钼锑抗比色法测定土壤速效磷含量,采用1.0 mol/L NH4OAc浸提-火焰光度法测定土壤速效钾含量,采用重铬酸钾氧化外加热法测定土壤有机质含量。供试土壤类型为砂姜黑土,基础肥力:碱解氮含量68.6 mg/kg,有效磷含量20.8 mg/kg,速效钾含量70.9 mg/kg,有机质含量11.1 g/kg。
1.2 试验设计
试验设CK(完全空白,不施肥)、K0(钾空白,不施钾肥)、K75(施K2O 75 kg/hm2)、K150(施K2O 150 kg/hm2)和K225(施K2O 225 kg/hm2),参照甘薯实际生产中钾肥上限用量,共5个处理。其中K0、K75、K150和K225处理均施氮75 kg/hm2作底肥。试验采用传统的起垄净作栽培方式,栽种密度为 49 500株/hm2,株距0.25 m,垄距0.8 m,小区面积50.4 m2(9 m×5.6 m),各处理重复3次。
1.3 测定项目与方法
分别于移栽后50、10、150 d的 09:00—11:00采用CIRAS-3便携式光合测定仪(Hansatech,USA)测定净光合速率(Pn)。用SPAD仪测定第5张功能叶的SPAD值;荧光参数采用FMS-2便携式脉冲调制式荧光仪(英国Hansatech公司生产)进行测定。
钾含量:先用H2SO4-H2O2消煮,然后使用火焰光度计进行测定。总淀粉和直链淀粉含量:将300 g甘薯洗净后切成小块,使用植株搅拌机搅拌至糊状,倒入100目纱袋,在0.5 L水中冲洗3次,并将冲洗液过100目筛,静置12 h;沉淀后的淀粉放于50 ℃烘箱中干燥24 h,随后研磨过100目筛,用酶法分析淀粉含量[14],重复3次。将纯化的直链淀粉作为标样,采用碘-碘化钾双波长比色法测定其含量。总淀粉含量与直链淀粉含量之差即支链淀粉含量。
各小区在甘薯收获期取20株记录茎叶鲜质量和块根鲜质量等,各处理随机选取5处9 m2核算产量。将茎叶切碎混匀,称取鲜样200 g左右于80 ℃烘至恒质量。将块根切成粒状并混匀后取150 g烘干测定干物质量。
1.4 计算公式
冠根比(T/R)=地上部生物量/地下部生物量;
淀粉产量(kg/hm2)=甘薯产量(kg/hm2)×干率(%)×淀粉含量(%);
钾积累量(mg/株)=甘薯生物量(g/株)×钾含量(%);
光系统Ⅱ实际光化学效率(ΦPSⅡ)=(Fm′-Fs)/Fm′;
光化学淬灭系数(qp)=(Fm′-Fs)/(Fm′-Fo′)。
式中:Fm′为光适应下最大荧光产量,Fs为稳态荧光产量,Fo′为光适应下最小荧光产量。
1.5 数据分析
用Microsoft Excel 2010预处理,经SPSS 18.0进行数据统计分析,采用SigmaPlot 12.5软件作图。
2 结果与分析
2.1 施钾量对甘薯生物量积累及冠根比的影响
由表1可知,与K0处理相比,各施钾处理均能提高甘薯各生长期地上和地下部生物量。移栽后50 d(发根结薯期),随着施钾量升高,冠根比(是反映甘薯干物质分配的重要指标)呈降低趋势,表明施钾有利于甘薯发根结薯期块根的早期膨大;移栽后100 d(薯蔓并长期),随着施钾量升高,冠根比呈升高趋势,表明施钾有利于此时期地上部源器官的建立,从而保证光合产物的供给;移栽后150 d(收获期),随着施钾量升高,冠根比整体上呈降低趋势,表明施钾有利于薯块膨大后期干物质从地上部向地下部运转,促进薯块膨大。
2.2 施钾量对甘薯产量的影响
由图1可知,与K0处理相比,2个甘薯品种的各施钾处理均能提高甘薯产量,且随着施钾量的增加,甘薯产量呈上升趋势。相同钾肥用量处理条件下,商薯19甘薯产量均高于徐薯18,其中以施钾量225 kg/hm2处理最高,达49 500.5 kg/hm2。從增产效果来看,与K0处理相比,商薯19低钾处理(K75)增产效果不佳,增幅仅为5.9%,而高钾处理下(K225)增产幅度较为明显,高达23.5%; 与K0处理相比,徐薯18低钾处理(K75)表现出较好的增产效果,幅度为16.5%,但增产效果随着施钾量的提高而下降,中钾处理(K150)增幅为25.7%,但高钾处理下(K225)增幅仅为28.8%,不同施钾处理间表现出明显的报酬递减现象。
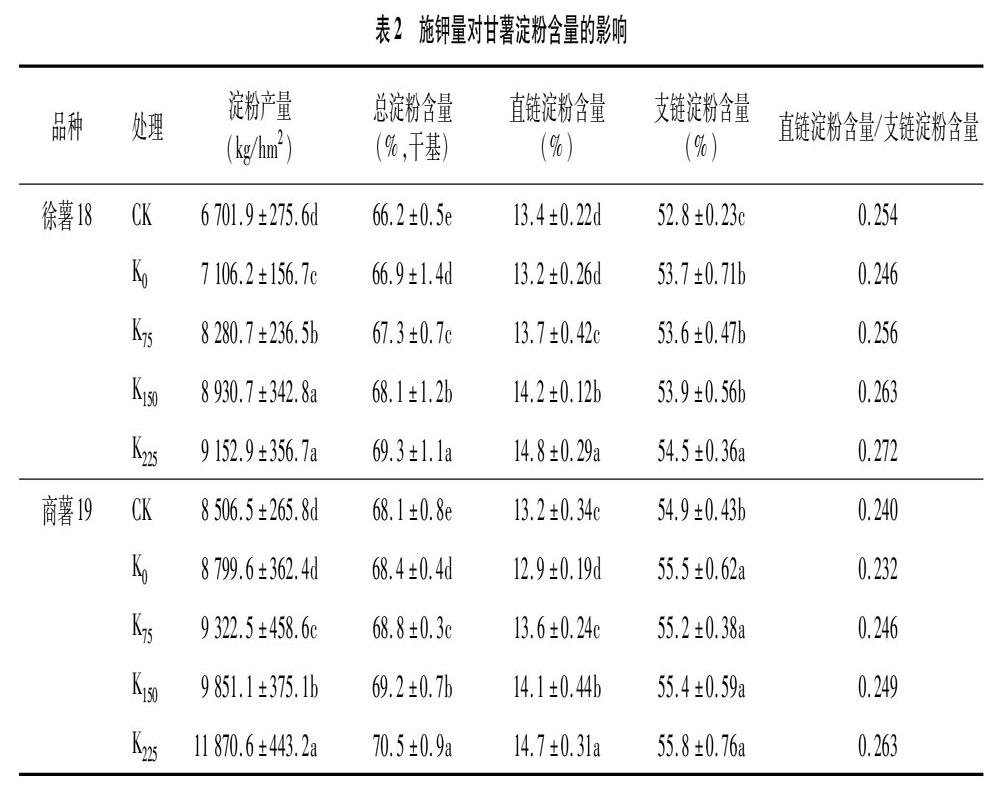
2.3 施钾量对甘薯淀粉含量的影响
由表2可知,与K0处理相比,徐薯18的3个施钾处理的淀粉产量分别提高了16.5%、25.7%和 28.8%;商薯19的3个施钾处理的淀粉产量分别提高了5.9%、11.9%和34.9%。随着施钾量的升高,2个甘薯品种的总淀粉含量呈显著升高趋势,其中徐薯18各施钾处理分别较K0处理提高0.4、1.2、2.4百分点,商薯19各施钾处理分别较K0处理提高0.4、0.8、2.1百分点。增加钾肥施用量能显著提高甘薯直连淀粉含量,进而提高甘薯直链淀粉含量/支链淀粉含量。在同一钾肥施用量处理下,徐薯18的总淀粉含量要低于商薯19,但直链淀粉含量呈相反趋势。
2.4 施钾量对甘薯钾积累量的影响
由表3可知,与K0处理相比,2个甘薯品种的各施钾处理均能提高甘薯钾积累量。随着施钾量的升高,甘薯各生长期的钾积累量呈上升趋势,且各施钾处理间差异显著,表明施钾能促进甘薯对钾素的吸收,进而增加甘薯钾素积累量。在同一钾肥施用量处理和生长期条件下,商薯19的钾素积累量整体上高于徐薯18。
2.5 施钾量对甘薯叶绿素含量及净光合速率的影响
由表4可知,与K0处理相比,2个甘薯品种的各施钾处理均能提高叶片SPAD值(叶片的叶绿素相对含量)和净光合速率(Pn)。移栽后100 d,甘薯由薯蔓并长期转入薯块膨大期,随着施钾量升高,叶片SPAD值和净光合速率整体上呈显著上升趋势, 均以施钾225 kg/hm2处理最高, 表明施钾能加速光合产物在叶片中的合成。移栽后150 d,甘薯地上部已衰老脱落,此时期随着施钾量升高,2个甘薯叶片SPAD值和净光合速率呈上升趋势(P<0.05),表明增施钾肥能延缓叶片衰老,叶片较高的叶绿素含量和净光合速率能保证块根中淀粉合成所需光合产物的持续供给。在相同施钾条件下,商薯19的SPAD值(除K225处理)和Pn均低于徐薯18。
2.6 施钾量对甘薯叶绿素荧光特性的影响
经叶片原初反应合成的光合产物由茎蔓运输到块根后转化并合成淀粉,因此PSⅡ是调控淀粉合成的关键因素之一。由表5可知,移栽后100 d(薯蔓并长期),与K0处理相比,2个甘薯品种各施钾处理均提高了叶片最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率(ΦPSⅡ)、qp、电子传递速率(ETR),这表明增施钾肥能提高薯块膨大期甘薯叶片PSⅡ反应中心的开放程度,通过提高电子传递速率、减少热耗散等一系列途径来加强对光能的利用,进而加速淀粉合成所需光合产物的积累。移栽后150 d(收获期)甘薯叶片已衰老脱落,各参数较移栽 100 d 时显著降低, 表明甘薯老叶光合结构已遭到破坏,PSⅡ反应中心可能部分关闭,PSⅡ光合作用活力受到抑制,光合电子传递受阻。此时期随着施钾量的增加,2个甘薯品种Fv/Fm、ΦPSⅡ、qp和ETR均呈升高趋势。说明增施钾肥能减缓光合结构受损程度,延缓叶片衰老,延长光合作用持续时间,保证薯块中淀粉合成所需光合产物的持续供给。
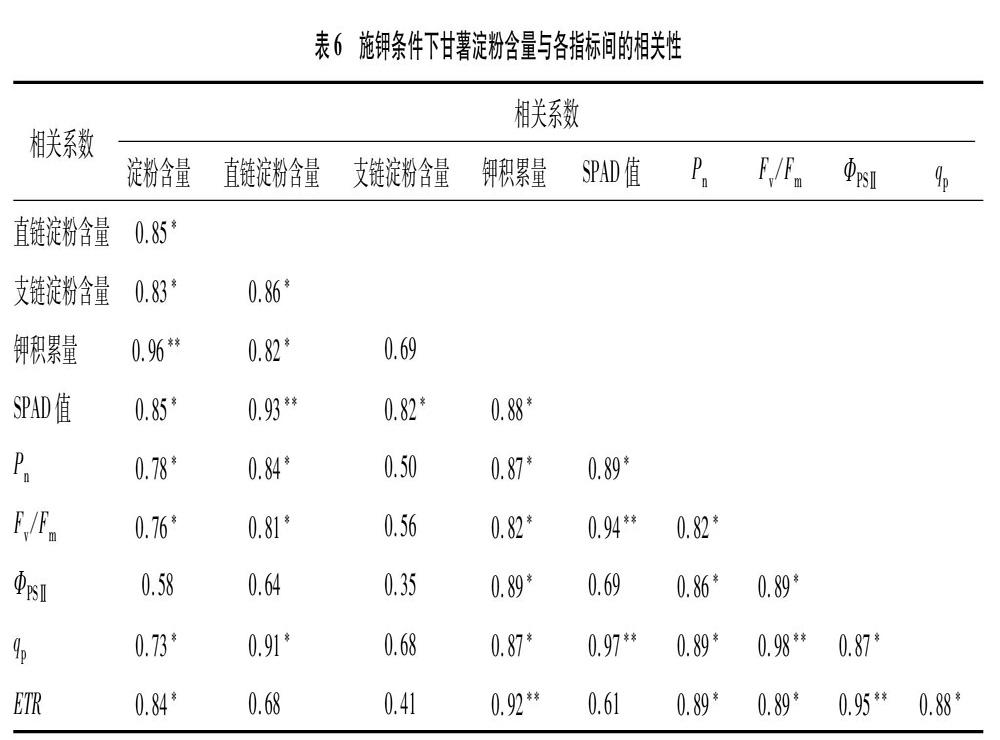
2.7 相关性分析
由表6可知,甘薯淀粉含量与SPAD值、Pn、Fv/Fm、qp、ETR呈显著正相关,与钾积累量呈极显著正相关(r=0.96,P<0.01)。钾积累量与直连淀粉含量、SPAD值、Pn、Fv/Fm、ΦPSⅡ、qp、ETR呈显著或极显著正相关。Pn与SPAD值、Fv/Fm、ΦPSⅡ、qp、ETR呈显著正相关。
3 讨论
3.1 施钾对甘薯淀粉含量的影响
淀粉含量是制約淀粉型甘薯产量的重要因素之一。淀粉含量不仅受到水分、温度和光照的影响[15],同时还受养分的调控[16]。本研究发现,与K0处理相比,徐薯18的3个施钾处理的淀粉产量分别提高了16.5%、25.7%和28.8%;商薯19的3个施钾处理的淀粉产量分别提高了5.9%、11.9%和34.9%。随着施钾量的升高,2个甘薯品种的总淀粉含量呈显著升高趋势,与K0处理相比,其中徐薯18各施钾处理分别提高0.4、1.2、2.4百分点,商薯19各施钾处理分别提高0.4、0.8、2.1百分点。这与Zhang等的研究结果[10,17]一致。本研究还发现,增加钾肥施用量对2个甘薯品种支链淀粉含量影响较小,主要显著提高甘薯直链淀粉含量,进而提高甘薯直链淀粉含量/支链淀粉含量,这与柳洪鹃等的研究结果[18]类似。淀粉的生物合成主要经过以下3个过程:(1)腺苷二磷酸葡萄糖(ADPG)的产生;(2)在游离淀粉合成酶(SSS)催化条件下合成支链淀粉;(3)在束缚态淀粉合成酶(GBSS)催化下完成直链淀粉的合成。
3.2 施钾对甘薯淀粉含量和光合特征相关性调控
甘薯块根膨大是光合作用形成的糖在淀粉合成酶的作用下转化为淀粉并在块根中不断累积的过程[18],因此,光合特性的变化是影响淀粉合成的第一步。本研究发现,随着施钾量的增加,各时期甘薯叶片SPAD值和Pn呈上升趋势。增施钾肥能提高薯块膨大期叶片叶绿素含量和Pn,利于光合产物积累;同时,充足的供钾使收获期叶绿素含量和Pn保持较高水平,延缓叶片衰老,延长光合作用持续时间,保证薯块膨大所需光合产物的持续供给[19-21]。随着施钾量的升高,Fv/Fm和ΦPSⅡ上升,表明在试验范围内增施钾肥能提高光能的利用能力,与此同时ETR显著升高[22]。随着施钾量的提高,叶片的qp显著升高,增施钾肥能够促进叶片光合产物的合成[23-25],使植物光合能力提升。较高的光合能力为甘薯淀粉合成提供了基础。
通过相关性分析发现,甘薯淀粉含量与钾积累量、光合特性存在显著的线性关系,甘薯淀粉含量与SPAD值、Pn、Fv/Fm、qp、ETR呈显著正相关,与钾积累量呈极显著正相关(r=0.96)。钾积累量与直连淀粉含量、SPAD值、Pn、Fv/Fm、ΦPSⅡ、qp、ETR呈显著或极显著正相关。可见,增施钾肥增加了甘薯植株内钾含量,从而提升了叶绿素含量、光系统Ⅱ活性和净光合速率,增加了块根直链淀粉含量,进而提高了总淀粉含量与淀粉产量。
3.3 不同品种甘薯淀粉含量对钾的响应规律
甘薯总淀粉含量、直链淀粉和支链淀粉含量主要受WX基因控制[26]。本研究发现,不同品种甘薯的总淀粉含量及构成成分不同。此外,环境条件能够影响总淀粉含量,但品种基因型是决定直链淀粉含量的主要因素。本试验发现,不同甘薯品种总淀粉含量与构成成分存在差异。在相同施钾用量处理下,徐薯18总淀粉含量和淀粉产量均低于商薯19。从构成成分角度看,2个甘薯品种的直链淀粉含量差异不显著,商薯19支链淀粉含量均显著高于徐薯18,可见支链淀粉含量是引起2个甘薯品种淀粉含量存在差异的主要原因。徐薯18施钾量超过 150 kg/hm2 时,产量和淀粉产量不再显著增加,而商薯19在施钾量为225 kg/hm2时,产量和淀粉产量达到最大值。
4 结论
增施钾肥能够增加甘薯植株内钾含量,从而提升其叶绿素含量、光系统Ⅱ活性和净光合速率,增加块根直链淀粉含量,进而提高总淀粉含量与淀粉产量。参照甘薯产量和淀粉含量因素,徐薯18推荐中量钾肥用量(150 kg/hm2),商薯19推荐高量钾肥用量(225 kg/hm2),相应钾肥用量能增加甘薯的产量和总淀粉含量,具有显著的经济价值和推广意义。
参考文献:
[1]易中懿,汪 翔,徐雪高,等. 品种创新与甘薯产业发展[J]. 江苏农业学报,2018,34(6):206-214.
[2]唐忠厚,张爱君,陈晓光,等. 低钾胁迫对甘薯块根淀粉理化特性的影响及其基因型差异[J]. 中国农业科学,2017,50(3):513-525.
[3]Trung P T B,Ngoc L B B,Hoa P N,et al. Impact of heat-moisture and annealing treatments on physicochemical properties and digestibility of starches from different colored sweet potato varieties[J]. International Journal of Biological Macromolecules,2017,105:1071-1078.
[4]Cui S,Li M,Zhang S,Liu J,et al. Physicochemical properties of maize and sweet potato starches in the presence of cellulose nanocrystals[J]. Food Hydrocolloids,2018,77:220-227.
[5]Akin-Ajani O D,Itiola O A,Odeku O A.Evaluation of the disintegrant properties of native and modified forms of fonio and sweet potato starches[J]. Starch,2016,68(1/2):169-174.
[6]Zhou D N,Zhang B,Chen B,et al. Effects of oligosaccharides on pasting,thermal and rheological properties of sweet potato starch[J]. Food Chemistry,2017,230:516-523.
[7]Tang Z H,Zhang A J,Wei M,et al. Physiological response to potassium deficiency in three sweet potato (Ipomoea batatas[L.]Lam.) genotypes differing in potassium utilization efficiency[J]. Acta Physiologiae Plantarum,2015,37(8):184.
[8]Kitahara K,Nakamura Y,Otani M,et al. Carbohydrate components in sweetpotato storage roots:their diversities and genetic improvement[J]. Breeding Science,2017,67(1):62-72.
[9]陈晓光,史春余,李洪民,等. 施钾时期对食用甘薯光合特性和块根淀粉积累的影响[J]. 应用生态学报,2013,24(3):759-763.
[10]Zhang K,Wu Z D,Tang D B,et al. Comparative transcriptome analysis reveals critical function of sucrose metabolism related-enzymes in starch accumulation in the storage root of sweet potato[J]. Frontiers in Plant Science,2017,8:914.
[11]Yooyongwech S,Samphumphuang T,Tisarum R,et al. Arbuscular mycorrhizal fungi (AMF) improved water deficit tolerance in two different sweet potato genotypes involves osmotic adjustments via soluble sugar and free proline[J]. Scientia Horticulturae,2016,198:107-117.
[12]Wang J D,Zhu G P,Dong Y,et al. Potassium starvation affects biomass partitioning and sink-source responses in three sweet potato genotypes with contrasting potassium-use efficiency[J]. Crop and Pasture Science,2018,69(5):506-514.
[13]張海燕,解备涛,段文学,等. 不同时期干旱胁迫对甘薯光合效率和耗水特性的影响[J]. 应用生态学报,2018,29(6):1850-1943.
[14]Kays S J. The physiology of yield in the sweet potato[M]// Sweet Potato Products. USA:CRC Press,2018.
[15]张飒琪,陈 杰,张 辉,等. 不同土壤水分条件下施氮对甘薯干物质积累及块根品质的影响[J]. 江苏农业学报,2013,29(3):533-539.
[16]柳洪鹃,姚海兰,史春余,等. 施钾时期对甘薯济徐23块根淀粉积累与品质的影响及酶学生理机制[J]. 中国农业科学,2014,47(1):43-52.
[17]Katayama K,Nishinaka M,Nakamura Y,et al. New sweet potato lines have high amylose and resistant starch contents[J]. Starch,2019,71(3/4):1800180.
[18]柳洪鹃,史春余,张立明,等. 钾素对食用型甘薯糖代谢相关酶活性的影响[J]. 植物营养与肥料学报,2012,18(3):724-732.
[19]Battie-Laclau P,Laclau J P,Beri C,et al. Photosynthetic and anatomical responses of Eucalyptus grandis leaves to potassium and sodium supply in a field experiment[J]. Plant,Cell & Environment,2014,37(1):70-81.
[20]Gupta R,Min C W,Wang Y,et al. Expect the unexpected enrichment of “hidden proteome” of seeds and tubers by depletion of storage proteins[J]. Frontiers in Plant Science,2016,7:761.
[21]Wijewardana C,Reddy K R,Shankle M W,et al. Low and high-temperature effects on sweetpotato storage root initiation and early transplant establishment[J]. Scientia Horticulturae,2018,240:38-48.
[22]Khanghahi M Y,Pirdashti H,Rahimian H,et al. Leaf photosynthetic characteristics and photosystem Ⅱ photochemistry of rice (Oryza sativa L.) under potassium-solubilizing bacteria inoculation[J]. Photosynthetica,2019,57(2):500-511.
[23]Ruiz C A S,van den Berg C,Wijffels R H,et al. Rubisco separation using biocompatible aqueous two-phase systems[J]. Separation and Purification Technology,2018,196:254-261.
[24]Dana S,Herdean A,Lundin B,et al. Retracted:each of the chloroplast potassium efflux antiporters affects photosynthesis and growth of fully developed arabidopsis rosettes under short-day photoperiod[J]. Physiologia Plantarum,2016,158(4):483-491.
[25]Liu W H,Zhan X W,Xing F L,et al. Effect on the grown of cotton seedlings and efficiency of potassium under different levels of light and applying potassium[J]. Cotton Sciences,2015(4):3.
[26]Xia J,Zhu D,Wang R,et al. Crop resistant starch and genetic improvement:a review of recent advances[J]. Theoretical and Applied Genetics,2018,131(12):2495-2511.
