基于Cytb基因的秆野螟属(Ostrinia)昆虫遗传多样性及其系统发育研究
2020-07-17杨瑞生顾羽健白树雄何康来王振营
杨瑞生 ,顾羽健 ,白树雄 ,何康来 ,王振营
(1.沈阳农业大学 生物科学技术学院/辽宁省昆虫资源工程技术研究中心,沈阳 110161;2.中国农业科学院 植物保护研究所/植物病虫害生物学国家重点实验室,北京 100094)
秆野螟属(Ostrinia)昆虫在全世界已报道21种,我国记录16种,是秆野螟属昆虫报道记录最多的国家[1-3],其中欧洲玉米螟(O.nubilalis)和亚洲玉米螟(O.furnacalis)是该属昆虫的优势种,为重要的世界性农业害虫。秆野螟属昆虫为一类小型昆虫,其表型分类及系统进化关系复杂[4-5]。秆野螟属昆虫的分类研究最早可追溯到1825年,HÜBNER将现在的酸模螟(O.palustralis)从Pyralis属归到Ostrinia属[1]。 1970年,MUTURRA 和MUNROE[1]依据成虫的形态特征对世界秆野螟属昆虫进行了整理和修订,将该属分为3个分支,并认为O.penitalis是秆野螟属中最原始的一个种。20世纪80年代至21世纪初,科研人员对秆野螟属的研究主要集中在发生规律、分类性状以及新种记录等方面[2,6-10]。秆野螟属昆虫种间成虫及幼虫形态相似度高,寄主及生态区重叠严重,其分类特征主要集中在雄性成虫外生殖器结构及中足胫节膨大与否等表型性状方面[11-14]。随着分子生物学及生物技术的发展,秆野螟属昆虫系统分类与进化研究逐渐展开,姜仲雪和孟祥锋[15]利用同工酶技术对秆野螟属5种近缘种的系统分类进行了研究;KIM等[16]利用线粒体DNA COⅡ基因序列探讨了分布于日本及北美的亚洲玉米螟等8种秆野螟属昆虫的系统进化关系;TABATA和ISHIKAWA[17]探讨了豆秆野螟(O.scapulalis)和刺菜螟(O.zealis)性信息素多态性的遗传学基础;OHNO等[18]分析了日本两种秆野螟属昆虫线粒体DNA COⅡ基因序列差异;杨瑞生等[5]利用线粒体DNA COⅠ基因探讨了我国秆野螟属昆虫系统进化关系。上述研究均表明秆野螟属昆虫亲缘关系较近,且种间界限不明显。
线粒体基因是目前应用于生物系统进化与发育研究最为广泛的分子标记之一。近年来,线粒体DNACytb基因结构与功能的研究表明该基因碱基序列稳定性高,进化速率适中[19],适合属间、种间甚至种内的系统进化关系研究[20],已经成为研究生物系统进化与分类鉴定研究的有力工具[21-23]。本研究探讨线粒体Cytb基因在秆野螟属昆虫系统进化研究中的可行性,以期为深入研究该属昆虫的物种分化与鉴定奠定理论基础。
1 材料与方法
1.1 供试昆虫
于国内不同地区采集秆野螟属昆虫的越冬幼虫,饲养于指形管内,室内化蛹、羽化,将成虫鉴定、分类后,于-20℃保存,备用。昆虫样品采集信息见表1。
1.2 主要试剂
PCR扩增引物由上海生工生物工程技术服务公司(Sangon)合成。
DNA提取试剂购自北京六合通经贸有限公司;PCR试剂购自宝生物(大连)工程有限公司;扩增和测序引物由上海生工生物工程技术服务公司合成;PCR产物回收试剂盒(E.Z.N.A.TMDNA Isolation System)由OMEGA公司提供。
1.3 方法
1.3.1 基因组DNA的提取 DNA提取参考杨瑞生等[5]进行,每个种群选取3个个体。基因组DNA在0.8%琼脂糖凝胶上定性分析,利用NanoDrop ND1000紫外分光光度仪定量分析,供PCR扩增参考。
1.3.2 PCR引物设计与目的基因扩增 根据GenBank数据库欧洲玉米螟mtDNA全序列(AF467260)设计秆野螟属 Cyt b PCR 引物,Cytb-O-F:5′-GGTCCTTTACGATCTTCAT-3′,Cytb-O-R:5′-GAGAATTAGTTTCTGGGTTT-3′。
PCR 反应体系 50μL: 模板 DNA 2μL,dNTPs (20mmol·L-1)2μL,Taq 酶 (2.5U)0.5μL,10×buffer2.5μL,Mg2+(25mmol·L-1)2μL,引物各 2μL,ddH2O 37μL。 PCR 程序:94℃预变性 3min,接下来 35 个循环,每个循环包括 95℃变性 30s,50℃退火 45s,72℃延伸 1min,循环结束后,72℃延伸 7min。
1.3.3 PCR产物回收及测序 扩增产物利用DNA纯化试剂盒进行回收,样品由上海生工(Sangon)公司测序,测序引物同扩增引物。
1.3.4 基因序列分析 利用Chromas2.5.1软件将所测序列进行校正,将所得基因序列进行BLAST搜索,确认是否为目的基因。利用DNAMAN 8.0软件进行碱基组成和碱基偏好性(Skewness)分析,AT Skewness=(A-T)/(A+T),GC Skewness=(G-C)/(G+C)。 利用 PAUP 4.0[24]和 MEGA 7.0[25]软件包分析各物种间Cytb基因的碱基变异和遗传距离(p-distance)等。以近缘属稻纵卷叶螟Cytb基因(JN246082.1)为外群,分别采用最大似然法(ML)、贝叶斯法(BI)和最大简约法(MP)构建分子进化关系,并抽样1000次进行Bootstrap自检验[26]。
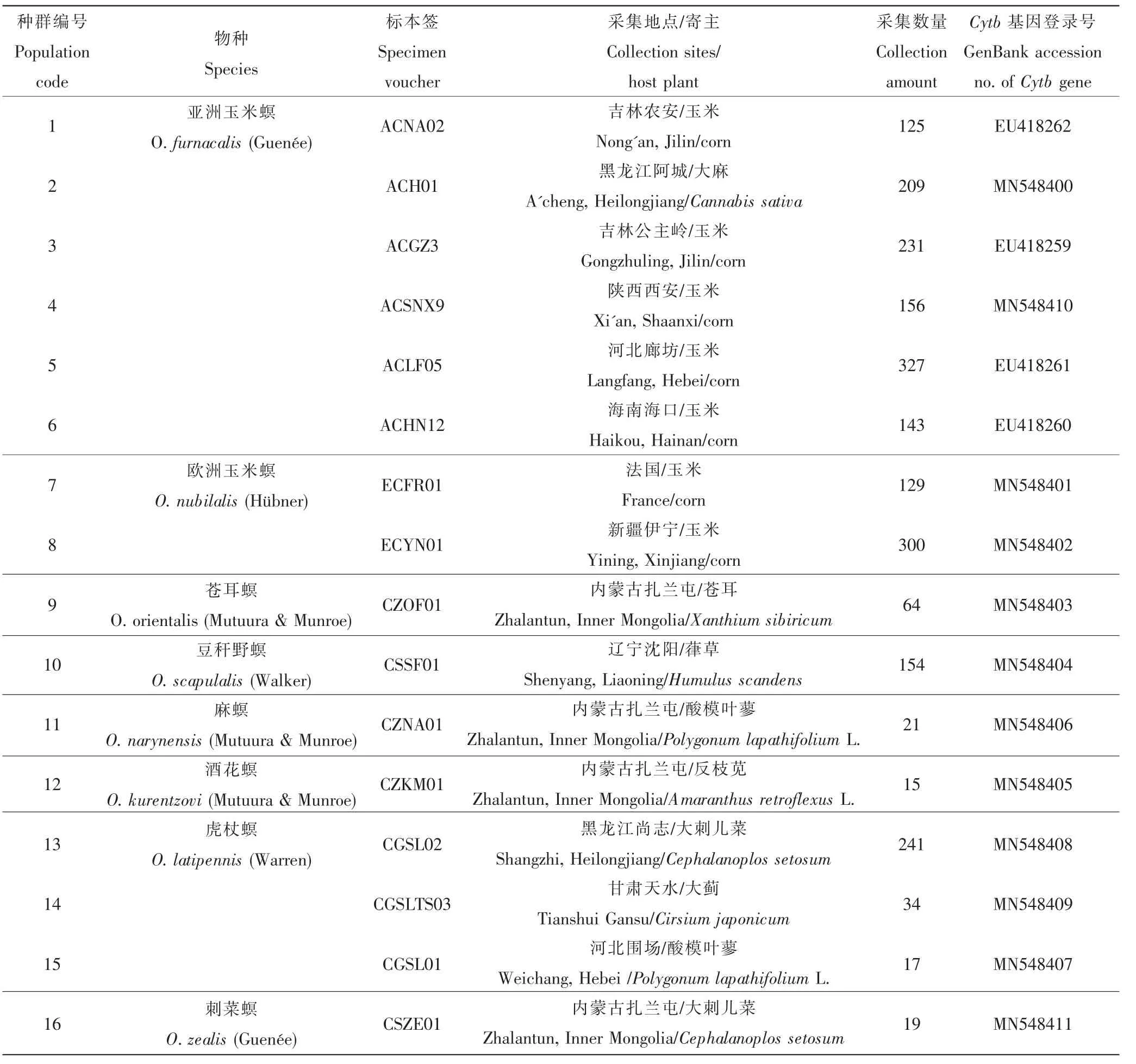
表1 秆野螟属昆虫样本采集信息及Cytb基因GenBank登录号Table 1 Data of Ostrinia spp.and GenBank accession numbers of Cytb genes
2 结果与分析
2.1 Cytb基因的确定
PCR结果表明,在1000~2000bp之间靠近1000bp的位置,所有样本均能成功扩增出单一特异性条带,分子量大小约1300bp,与预期结果一致(图1)。将所得序列在NCBI网站(www.ncbi.nlm.gov/)上进行相似性搜索,结果显示,得到的序列包括完整的Cytb基因和侧翼的部分基因序列。利用ClustalX 1.8软件[27]将所得到的序列与欧洲玉米螟线粒体DNA全序列(AF467260)进行比对,删除目的基因5'和3'两端的多余序列,得到秆野螟属Cytb基因的完整序列,目前,8个物种所有样品的Cytb基因序列已经提交到NCBIGenBank数据库(登录号见表1)。
2.2 秆野螟属Cytb基因碱基组成及其变异分析
秆野螟属昆虫Cytb基因序列全长1146bp,起始密码子为ATA,以密码子TAA终止。Cytb基因与5'端的ND6基因间有2个碱基的间隔区,与3'端的tRNA-Ser基因间无碱基间隔。8种螟虫Cytb基因1 146个核苷酸位点中没有碱基插入或缺失现象,其中有1051个保守性位点,95个可变性位点,变异率为8.29%,包括88个简约信息位点和7个单突变位点。Cytb基因碱基T、A、G和C的平均含量分别为41.1%、35.4%、10.1%和13.4%,AT和CG含量分别为76.5%、23.5%,表现出明显的碱基使用偏向性,其中碱基T的使用偏向性较弱(AT平均偏倚度为-0.0745),碱基C使用偏向性较强(GC平均偏倚度为-0.1441)。就秆野螟属不同昆虫种类而言,碱基T使用偏向性最强的为虎杖螟(AT偏倚度为-0.0861),其次是苍耳螟(AT偏倚度为-0.0731)、亚洲玉米螟(AT偏倚度为-0.0721)、酒花螟(AT偏倚度为-0.0719)、欧洲玉米螟(AT偏倚度为-0.0712)和麻螟(AT偏倚度为-0.0705),豆秆野螟和刺菜螟的T使用偏向性最弱,AT偏倚度均为-0.0693。碱基C使用偏向性最强的螟虫为亚洲玉米螟(GC偏倚度为-0.1550),其次是虎杖螟(GC偏倚度为-0.1474),刺菜螟的最弱(GC偏倚度为-0.1064)(表 2)。
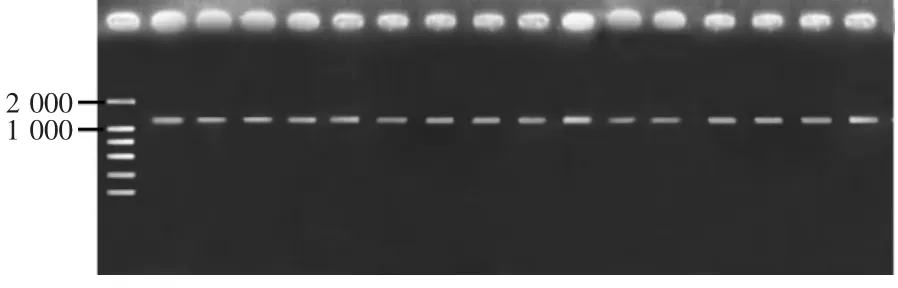
图1 秆野螟属Cytb基因扩增结果Figure 1 PCR results of Cytb genes from the genus Ostrinia
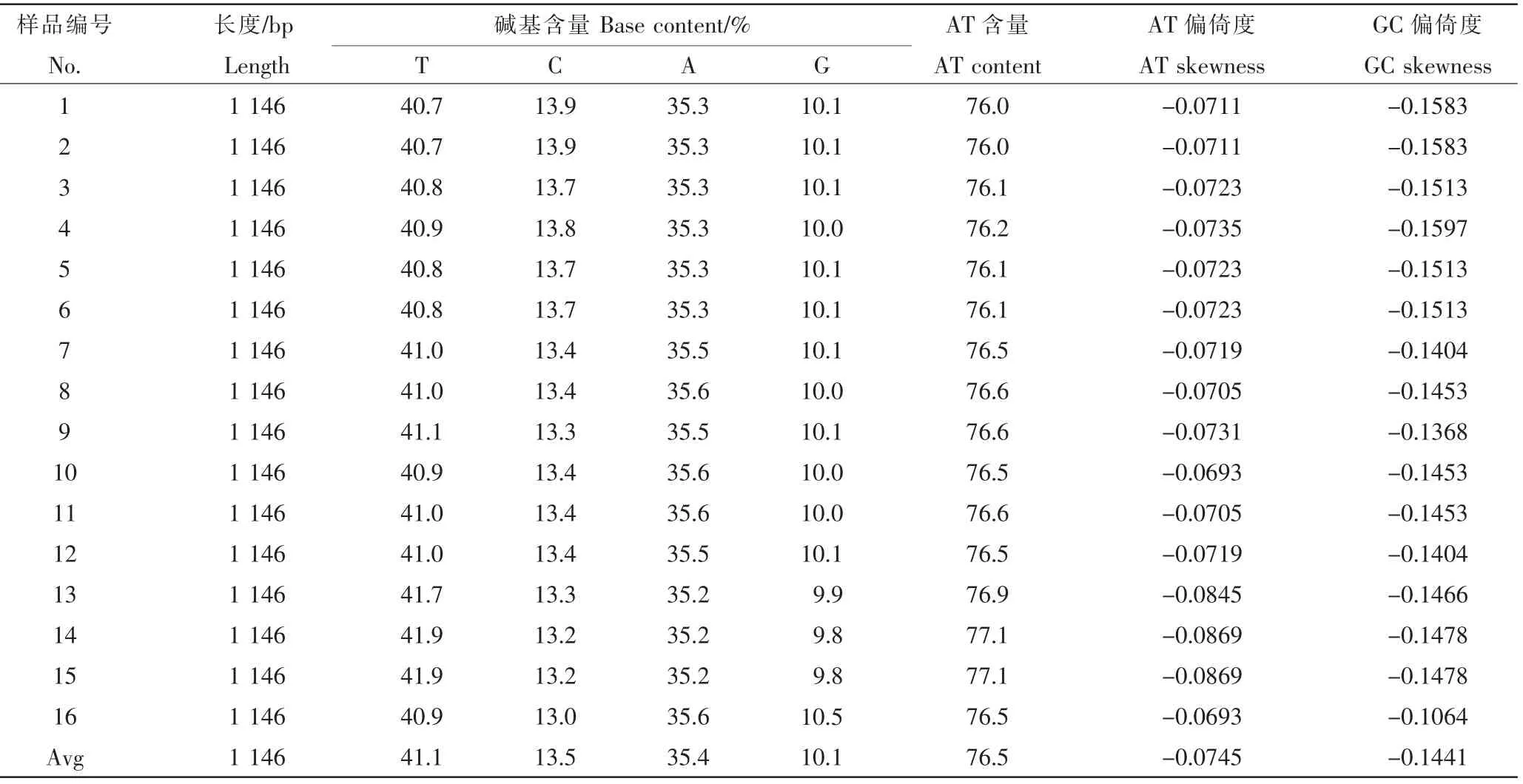
表2 秆野螟属昆虫Cytb基因碱基组成及碱基使用偏向性Table 2 Base composition and usage bias of Cytb genes from Genus Ostrinia
密码子不同位点的碱基组成,第3位点AT含量最高,达92.8%,第1和第2位点AT含量相近,分别为68.9%、67.7%(表3),Cytb基因的碱基组成和密码子碱基分布都表现出了较强的AT偏向性。
Cytb基因碱基替换模式中,碱基转换(Ts)数明显大于碱基颠换(Tv)数(平均R值为3.8),碱基置换占碱基变异总数的79.4%,碱基颠换占20.6%。其中,碱基置换主要发生在T←→C间,占碱基置换总数的81.5%,少数发生在A←→G间,占18.5%;碱基颠换主要发生在A←→T间,占颠换碱基的57.1%,少数发生在C←→A、T←→G之间,分别占总颠换碱基的28.6%和14.3%,没有C←→G间颠换的发生。Cytb基因密码子第3位点的碱基变异性最强(变异率为76.5%),其次是第1位点(变异率为20.6)%,第2位点碱基变异性最弱(变异率为2.9%),即第2位点碱基变异性最为保守(表4)。
秆野螟属昆虫线粒体DNACytb基因共编码氨基酸381个,其中15个氨基酸位点发生了变异 (变异率3.94%),包括简约信息位点10个(变异率2.63%)和单突变位点5个(变异率1.31%)。氨基酸组成上,异亮氨酸(Ileu)含量最高,占氨基酸总数的 15.61%,其次是亮氨酸(Leu)、苯丙氨酸(Phe)和天冬酰氨(Asn),含量依次为12.88%、8.89%、6.27%,含量最少的是半光氨酸(Cys),含0.79%(表5),存在明显的氨基酸组成偏向性。

表3 秆野螟属Cytb基因密码子不同位点的碱基组成Table 3 Base composition at defferent codon positions in Cytb genes from genus Ostrinia %
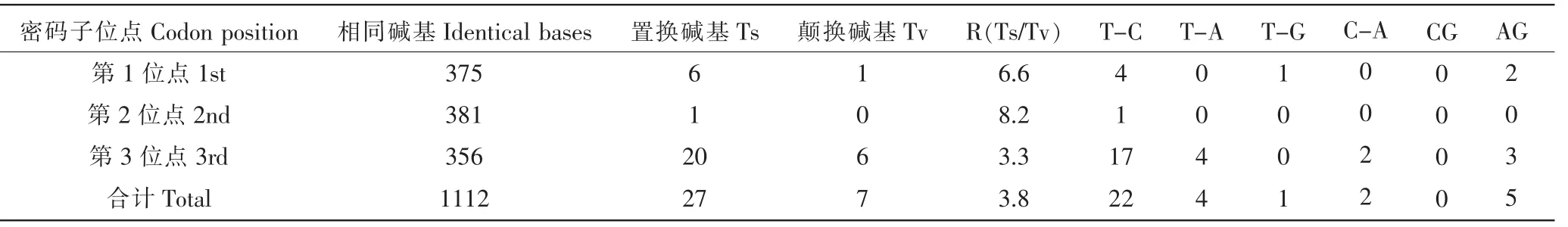
表4 秆野螟属Cytb基因碱基替换模式Table 4 Base substitution pattern in Cytb genes from genus Ostrinia
2.3 Cytb基因碱基差异及遗传距离分析
本研究中,秆野螟属昆虫种间遗传距离明显大于种内遗传距离。亚洲玉米螟6个不同地理种群间碱基差异位点有0~4个,遗传距离为0~0.0052,农安与尚志种群、廊坊与海南种群之间的Cytb碱基没有差异,遗传距离最远的发生在农安与西安种群、尚志与西安种群之间,均为0.0052;2个欧洲玉米螟种群间的碱基差异位点1个,遗传距离0.0009;虎杖螟的3个不同地理种群间碱基差异位点有0~1个,遗传距离在0~0.0017;秆野螟属种间差异位点有1~80个,遗传距离在0.0009~0.0698之间,遗传距离最小的发生在麻螟与酒花螟、苍耳螟与酒花螟之间,均为0.0009,最大的遗传距离发生在亚洲玉米螟西安种群与虎杖螟尚志种群之间,遗传距离为0.0698(表6)。

表5 秆野螟属线粒体Cytb氨基酸组成Table 5 Amino acid composition in Cytb genes of the genus Ostrinia
2.4 Cytb基因系统发育信号检验
以两两样本之间的遗传距离(p-distance)作为横坐标,碱基替换为纵坐标,绘制碱基替换模式散点图,进行秆野螟属昆虫Cytb基因序列碱基替换饱和度分析和系统发育信号检验(图2)。结果表明,绝大多数样本间的碱基置换数Ts高于碱基颠换Tv。碱基置换与遗传距离之间存在着明显的线性关系,即随着遗传距离的增大,碱基置换数远未达到饱和;而随着最初遗传距离的增大,碱基颠换也逐渐增大,当遗传距离达到0.05时,碱基颠换的变化逐渐趋于稳定,呈现饱和状态。受碱基置换数量远高于颠换的影响,碱基置换和颠换之和,即碱基替换与碱基置换的变化趋势一样,随着遗传距离的增大,也呈现出明显的线性关系,秆野螟属昆虫Cytb基因随着遗传距离的增大,碱基替换远未达到饱和,仍然处于线性上升的趋势。

表6 秆野螟属Cytb基因遗传距离(p-Distance)及碱基差异(上三角为碱基置换/碱基颠换;下三角为遗传距离)Table 6 Genetic distances (p-Distance) and base divergence of Cytb gene among Ostrinia (upper triangle:Ts/Tv;lower triangle:genetic distance)
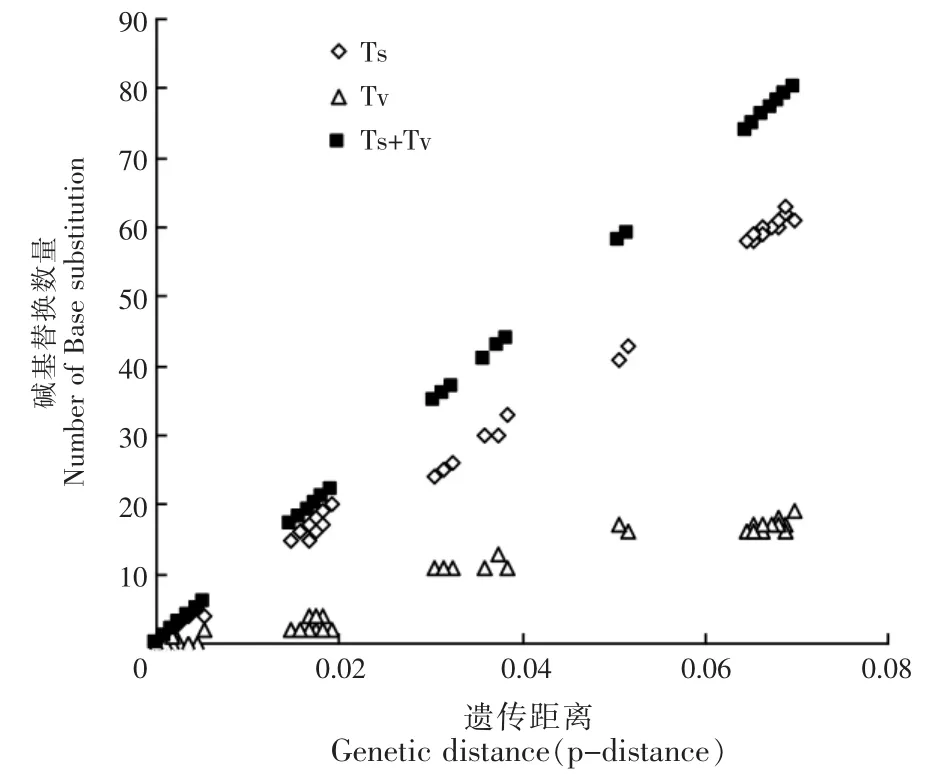
图2 秆野螟属昆虫Cytb基因碱基饱和度分析Figure 2 Saturation analysis of base substitution of Cytb genes from Ostrinia
2.5 Cytb基因分子系统树的构建
分别利用ML、BI和MP法,Bootstrap1000次自检,对秆野螟属昆虫线粒体DNACytb基因序列进行比较分析,得到了相应的系统进化树,进化树各分支的置信度在51~100之间(图3)。虽然种内及种间的拓扑结构稍有不同,但揭示的各物种间的系统进化关系却完全一致。结果表明,稻纵卷叶螟作为外群,单独形成一支,秆野螟属8种螟虫单独聚在一起,且分为二大支系,其中虎杖螟与其他7种螟虫亲缘关系最远,3个不同地理种群首先聚在一起,单独形成一个支系;亚洲玉米螟等7种螟虫的亲缘关系较近,聚在一起形成另一支系,其中分成2个分支,刺儿菜螟单独形成一个分支,而其他6种螟虫聚在一起形成另一分支。在该分支中,亚洲玉米螟不同地理种群首先聚在一起形成一个支系,酒花螟与麻螟互为姊妹种聚在一起,与苍耳螟、欧洲玉米螟和豆秆野螟亲缘关系较近聚在一起形成一个支系。
3 讨论与结论

图3 秆野螟属8种螟虫线粒体Cytb基因系统发育树Figure 3 Molecular phylogeny of 8 Ostrinia spp.based on Cytb genes
秆野螟属昆虫经典分类和系统发育研究主要依赖于雄性成虫外生殖器及中足胫节等部位的形态特征。根据上述性状,MUTUURA和MUNRROE[1]将世界秆野螟属昆虫分为3个支系。其中,O.penitalis由于雄性成虫外生殖器基环背部3分裂、抱器腹背部无刺,而单独分为一支;虎杖螟O.latipennis等9种螟虫由于雄性成虫外生殖器爪形突不分裂和抱器腹背部具刺等共同特征而聚为一个支系;亚洲玉米螟等10个种由于雄性成虫爪形突3分裂聚为一支,在这个支系中,又依据雄性成虫中足胫节大小,分为小胫节、中胫节和大胫节三个亚支(图4)。本研究中,中足胫节较大的豆秆野螟未能与胫节较小的亚洲玉米螟、欧洲玉米螟和苍耳螟分开,值得进一步探讨,其他物种间的拓扑关系则与MUTUURA和MUNRROE[1]的研究结果完全一致。
KIM等[16]利用线粒体COⅡ基因数据分析了日本和北美的8种秆野螟属昆虫的系统演化关系,结果显示,虎杖螟与酸模螟(O.palustralis)聚在一起单独形成一个支系,亚洲玉米螟等6种螟虫形成另一个支系,其中又根据中足胫节的大小分为大胫节和小胫节两个分支;杨瑞生等[5]以线粒体COⅠ基因序列为分子标记探讨了我国亚洲玉米螟等部分秆野螟属昆虫的系统进化关系。上述基于线粒体基因数据得到的秆野螟属昆虫进化关系与本研究结果完全一致。
昆虫线粒体Cytb基因的碱基组成与氨基酸序列的关系,尤其是高AT含量与氨基酸序列变异,甚至蛋白活性间有无相关性仍存在争议。CROZIER等[28]在研究线粒体基因碱基高AT含量与编码氨基酸关系时,认为AT高含量与氨基酸的编码种类与顺序无显著相关性;HUGALL等[29]则认为线粒体基因中碱基AT的高含量增加了氨基酸编码种类和序列的变化几率;DESALLE等[30]也认为高AT含量有提高碱基颠换发生的几率,进而导致密码子第3位点碱基颠换率的升高。本研究中,秆野螟属Cytb基因碱基组成差异较大,其中AT含量(76.5%)明显高于CG含量(23.5%)。就每个密码子的碱基位点来看,碱基组成也存在较大差异,第3位点AT含量显著高于GC含量,即线粒体Cytb基因存在明显的AT使用偏向性,这种高AT含量与Cytb编码氨基酸种类与顺序之间的相关性,需要进一步研究证明。
研究表明,线粒体基因突变中,碱基置换的发生频率较碱基颠换的发生频率更高[5,16,24]。本研究也证明了这一点,即秆野螟属昆虫Cytb基因的碱基置换数量随着样本间遗传距离的增加一直显著高于碱基颠换数量,且伴随物种的分化呈现出明显的线性上升趋势,使得整个碱基变异数量伴随物种间分化出现明显的线性关系。表明秆野螟属昆虫线粒体基因的进化与变异远未达到饱和,随着时间的推移和气候环境的变化,秆野螟属昆虫仍存在较大的进化潜力。
秆野螟属昆虫基于线粒体Cytb基因的遗传多样性较低,系统进化与发育关系较近,物种间Cytb基因进化不明显,但通过Cytb基因序列能够将秆野螟属昆虫各物种区分开,可以作为该属物种鉴定的辅助工具。
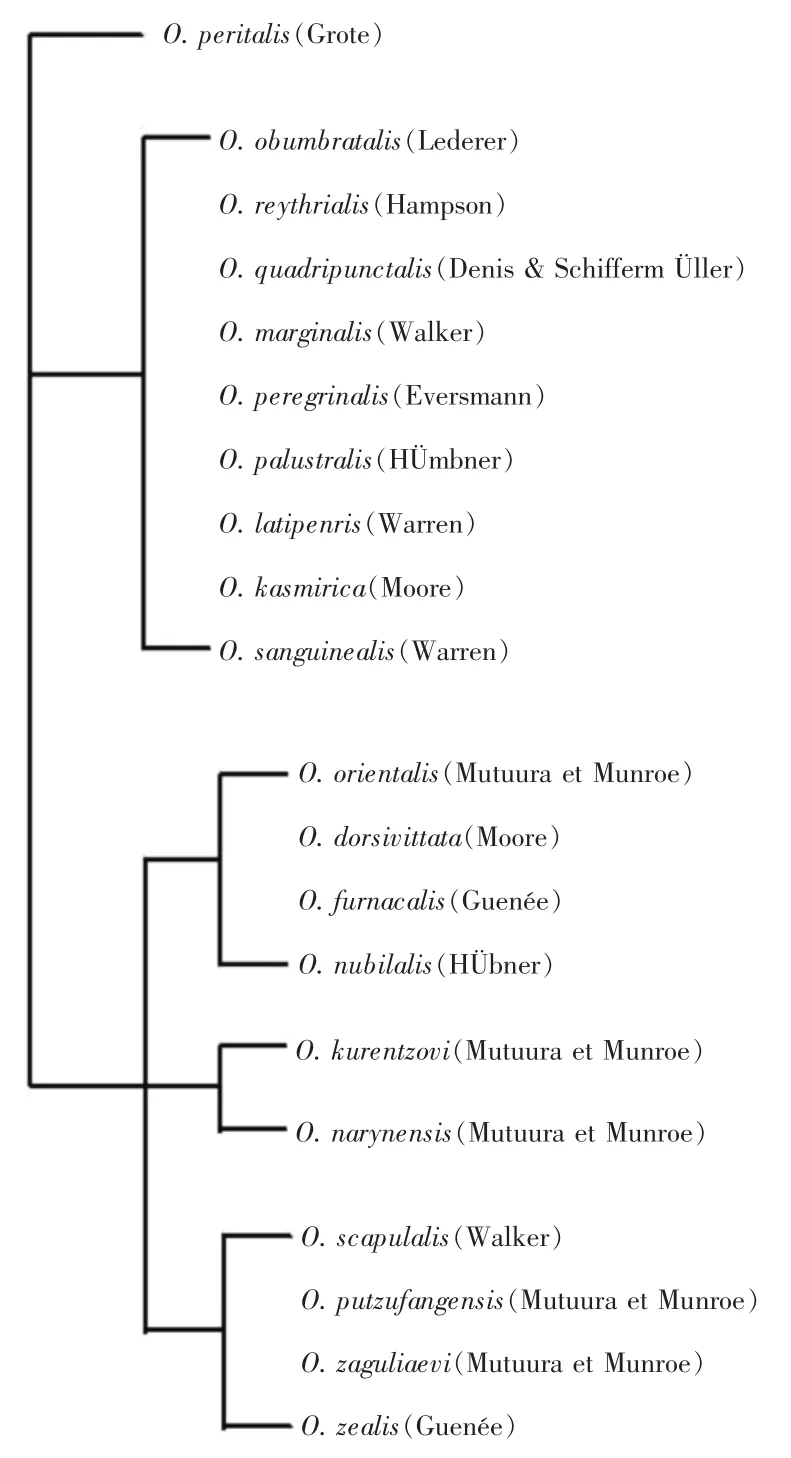
图4 秆野螟属昆虫的传统进化关系[1]Figure 4 Traditional taxa of the genus Ostrinia
