水分对敦煌阳关湿地芦苇叶片与土壤C、N、P生态化学计量特征的影响
2020-07-17包雅兰赵海燕
刘 冬,张 剑,包雅兰,赵海燕,陈 涛
西北师范大学地理与环境科学学院, 兰州 730070
在陆地生态系统中,营养元素相互作用过程中的平衡关系被称为生态化学计量学[1],生态化学计量学为研究碳(C)、氮(N)、磷(P)等多种元素在生态过程中的循环与耦合关系提供了一种重要的方法[2]。C是构成植物体内干物质的最主要元素,而N和P是各种蛋白质和遗传物质的重要组成元素[3],它们是地球生命化学的基础,也是连接生物体与自然资源的重要纽带[4]。植物通过根系从土壤中吸收生长发育所需的养分,叶片通过光合作用同化和积累C,又通过凋落物将所吸收养分反馈给土壤[5]。这是一个复杂的养分循环过程,不仅受植物自身生长型、生理特征和生活史等影响,而且受生长环境、群落组成及土壤理化因子等多方面影响。因此研究植物与土壤营养元素特征,对于揭示植物在不同生境下生长策略以及养分循环方式具有重要意义。
C、N、P化学计量比格局及其驱动因素一直是生态化学计量学的一个重要研究领域[6]。近30年来国内外学者对生态化学计量学的研究,不仅涉及到了生物学科的不同层次(分子、细胞、有机体、种群、生态系统)[6],而且还发展到了不同时空尺度(全球[7]、欧洲[8]、北美[9]、中国[10]、中国东部南北样带[11])。也不再聚焦于生态系统的某一个组分,而是综合考虑整个生态系统[12]。国内外学者对不同生态系统植物叶片-土壤生态化学计量学特征开展了大量研究。如俞月凤等[13]对喀斯特峰丛洼地不同森林类型植物和土壤C、N、P化学计量特征进行研究得出土壤C、N、P供应量对乔木叶片C、N、P含量影响不大,Yang等[1]对华北沙化地区不同植物群落间叶、根、凋落物和土壤的生态化学计量进行研究得出C∶N∶P化学计量可能引起土壤特性的变化,章广琦等[14]通过对黄土丘陵区刺槐与油松人工林生态系统生态化学计量特征进行研究得出不同层次植物与土壤养分特征间具有不同的关系,而对湿地生态系统植物与土壤生态化学计量特征及其相互作用的研究未受到较高的关注,尤其是对环境变化较为敏感的干旱区湿地。
干旱区湿地是江河源区重要的水源涵养地,在调节气候和遏制区域土地荒漠化方面有着不可替代的作用[15]。在干旱区,植物生长不仅受土壤养分限制,还受水分短缺限制。植物叶片容易受到生物与非生物因素的影响,对土壤水分有效性做出响应[16],随着干旱程度的加剧,叶氮、叶磷会维持在较高水平[17]。为此,本研究以土壤含水率为梯度,研究干旱区湿地芦苇(Phragmitesaustralis)叶片与土壤的生态化学计量特征及其关系,试图明晰:(1)不同水分梯度下的芦苇叶片和土壤C、N、P生态化学计量特征的差异性;(2)不同水分梯度下芦苇叶片与土壤C、N、P生态化学计量特征的关联性。以期为干旱区湿地资源的合理利用以及生态恢复提供科学依据。
1 材料与方法
1.1 研究区概况

图1 研究区地理位置及样地分布示意图Fig.1 The distribution map of observation sites
敦煌阳关湿地,位于距敦煌市70 km的敦煌阳关国家自然保护区东南4 km的黄水坝水库(渥洼池)一带(94°06′—94°09′E,39°51′—39°55′N)(图1),总面积11.06 km2,是中国西部荒漠区中较为罕见的特殊成因湿地生态系统。南与阿克塞哈萨克族自治县相邻,北与新疆维吾尔自治区接壤,西接库姆塔格沙漠。区内地势平坦,海拔高度在1150—1500 m之间,全年平均气温9.3℃,年均降雨量为36.9 mm,相对湿度43%,气候干燥少雨,夏季酷热,冬季寒冷,属于典型大陆性干旱气候,具有干旱区湿地生态系统到荒漠生态系统过渡的典型特征。区内有多处泉眼,量大且常年溢出,为湿地生态系统提供稳定补给水。主要土壤类型为沼泽土、草甸土和盐渍土,部分区域分布有风沙土和棕漠土。芦苇(Phragmitesaustralis)是研究区内的优势物种,伴生种有赖草(Leymussecalinus)、黑果枸杞(Lyciumruthenicum)、水麦冬(Triglochinpalustre)、盐角草(Salicorniaeuropaea)、鸦葱(Scorzoneraaustriaca)、海乳草(Glauxmaritima)等。
1.2 样地设置与采样
在2015年对研究区全面调查的基础上[15],2016年8月,根据代表性和典型性原则,通过聚类分析将18个样地分为低水分W1(n=6)、中水分W2(n=5)、高水分W3(n=7)三个梯度(高、中和低水分梯度土壤含水率分别为32.72%、22.27%和7.30%),在每个水分梯度上设置了5—7个30 m×30 m的样地,在每个样地内随机设置三个1 m×1 m的样方,共计54个样方。
在每个样方内根据芦苇根系生长状况,按0—10 cm,10—20 cm,20—40 cm,40—60 cm四个深度,沿样方对角线,分层钻取土样,去除杂质后放置在通风处阴干,研钵研磨后过0.25 mm和0.15 mm筛,密封低温保存;在每个样方内,采集相对完整的芦苇叶片若干带回实验室,在65℃条件下烘干至恒重,采用混合球磨仪(MM400,Retsch,Germany)磨碎后过0.15 mm筛,密封低温保存。
1.3 样品测定
采用电导法测定土壤盐分,采用烘干法测定土壤含水率,采用酸度计测定土壤pH,采用环刀法测定土壤容重;采用重铬酸钾-硫酸外加热法测定土壤有机碳(OC)和芦苇叶片C含量;采用开氏法测定土壤全氮(TN)和芦苇叶片N含量;采用钼锑抗比色法测定土壤全磷(TP)和叶片P含量[18]。
1.4 数据处理
利用SPSS 22.0(SPSS Inc. Chicago,USA)软件进行统计分析,用单因素方差分析检验不同水分梯度叶片和土壤C、N、P以及土壤理化性质的差异性,用Pearson相关分析方法分析叶片与土壤生态化学计量学之间的关系。
2 结果与分析
2.1 不同水分梯度芦苇叶片生态化学计量学特征
如图2所示,叶片C、N、P含量随土壤含水率的升高而降低。其中叶片C含量变化范围为365.36—442.43 mg/g,均值为408.52 mg/g,高水分梯度叶片C含量显著低于中、低水分梯度(P<0.05);叶片N含量变化范围为15.26—25.62 mg/g,均值为19.54 mg/g,不同水分梯度之间无显著差异(P>0.05);叶片P含量变化范围为0.50—1.65 mg/g,均值为1.03 mg/g,低水分梯度叶片P含量显著高于高、中水分梯度(P<0.05)。
叶片C/N、C/P、N/P随土壤含水率的升高而升高。C/N、C/P、N/P对应的均值分别为21.27、415.68和19.77,变化范围分别为20.84—21.70、344.21—455.81和16.76—21.60。高、中水分梯度叶片C/P、N/P显著高于低水分梯度(P<0.05),而叶片C/N不同水分梯度间差异不显著(P>0.05)。
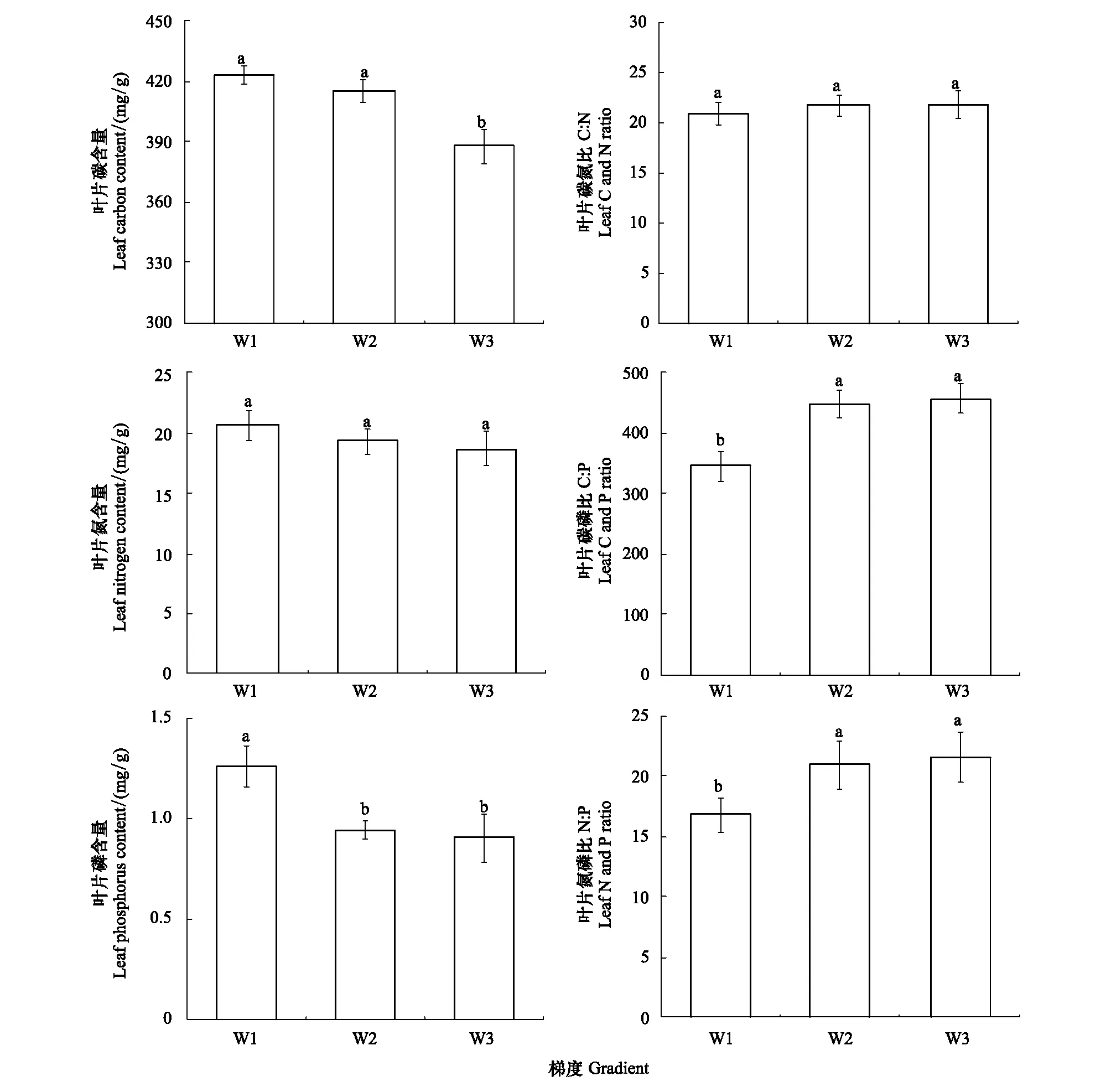
图2 不同水分梯度下芦苇叶片C、N、P化学计量特征(平均值±标准误差)Fig.2 Stoichiometric characteristics of C, N and P of reed leaves under different water gradients (mean±SE)W1:低水分梯度,Low moisture gradient;W2:中水分梯度,Medium moisture gradient;W3:高水分梯度,High moisture gradient;a、b、c表示不同水分梯度之间有显著差异(P<0.05)
2.2 不同水分梯度土壤生态化学计量学特征
如图3所示,土壤OC、TN、TP含量随土壤含水率的升高而升高。土壤OC含量变化范围为7.3—8.8 mg/g,均值为8.3 mg/g;土壤TN含量变化范围为0.41—0.78 mg/g,均值为0.58 mg/g,其中高水分梯度土壤TN含量显著高于中、低水分梯度(P<0.05);土壤TP含量变化范围为 0.40—0.45 mg/g,均值为0.42 mg/g。
土壤C/N、C/P、N/P的变化范围分别为10.4—19.0、16.4—22.9和0.95—1.45,均值分别为14.0、19.6和1.3。高水分梯度土壤N/P显著高于低水分梯度(P<0.05),高水分梯度土壤C/N显著低于中、低水分梯度(P<0.05)。
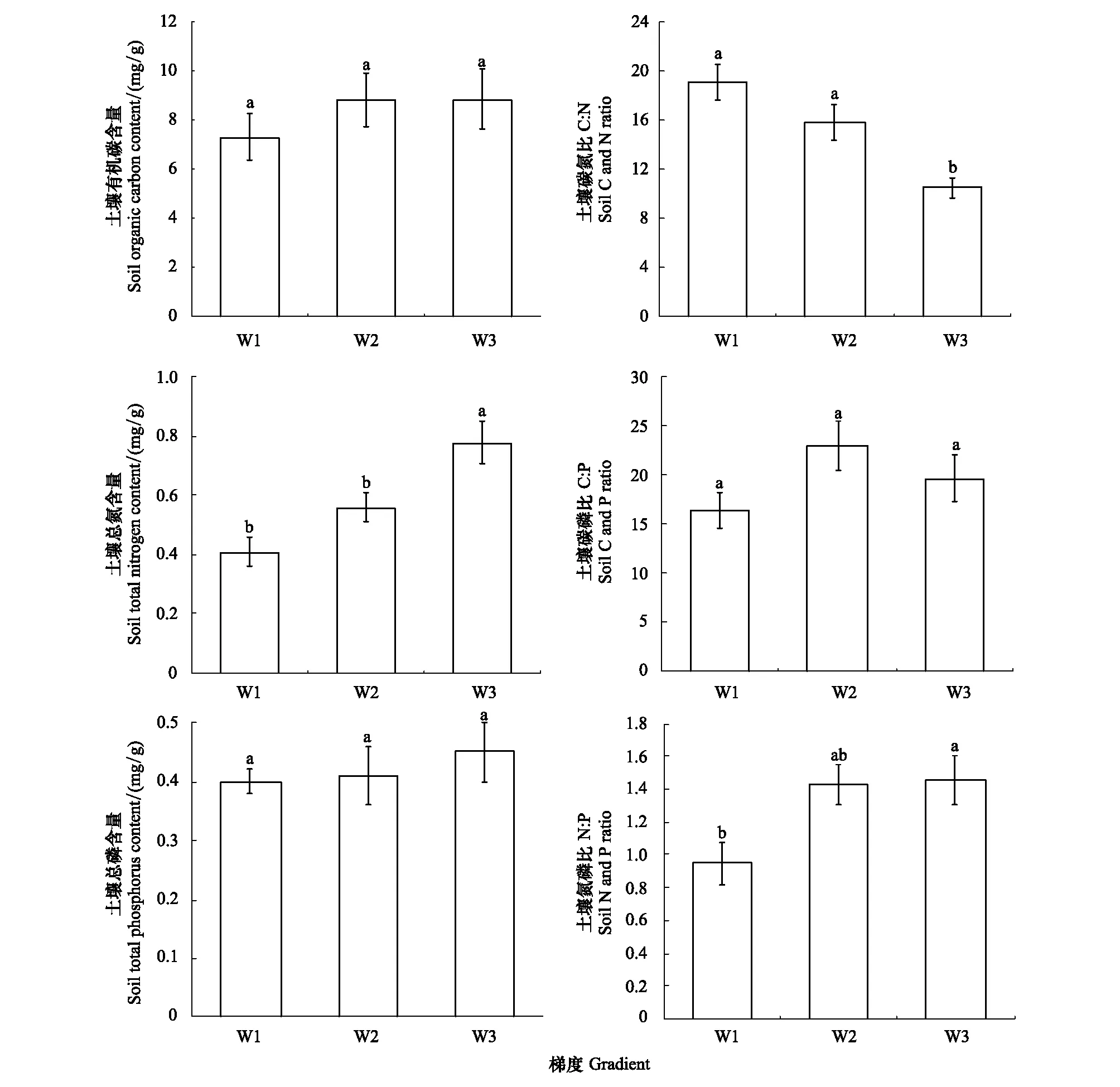
图3 不同水分梯度下土壤C、N、P化学计量特征(平均值±标准误差)Fig.3 Stoichiometric characteristics of soil C, N and P under different water gradients (mean±SE)
2.3 不同水分梯度土壤理化性质
由表1可知,从高水分梯度到低水分梯度,土壤水分显著下降了28.55%和72.89%;而土壤盐分显著上升了83.46%和59.54%。中水分梯度土壤pH显著高于高、低水分梯度,不同水分梯度土壤容重无显著差异。
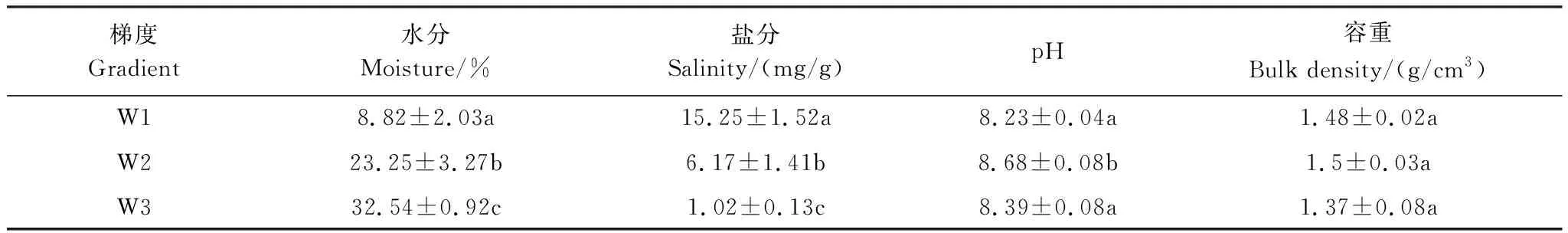
表1 不同水分梯度土壤理化性质表
2.4 不同水分梯度芦苇叶片和土壤化学计量学关系
由表2可知,高、中水分梯度芦苇叶片与土壤C、N、P化学计量特征无显著相关性(P>0.05)。在低水分梯度,土壤TN含量与叶片C、N含量显著负相关(P<0.05),与叶片C/N显著正相关;土壤TP含量与叶片C/P显著正相关(P<0.05);土壤C/N与叶片C含量显著正相关(P<0.05),与叶片N含量极显著正相关(P<0.01),与芦苇叶片C/N显著负相关;土壤N/P与叶片C、N含量显著负相关(P<0.05)。
3 讨论
3.1 不同水分梯度芦苇叶片C、N、P生态化学计量特征
高、中、低三个水分梯度叶片C、N、P平均含量(408.52、19.54 mg/g和1.03 mg/g)低于全球陆生植物叶片C、N、P平均水平(464、20.1 mg/g和1.99 mg/g)[7,19]、中国陆生植物叶片N、P平均水平(20.24 mg/g和1.21 mg/g)[10],主要因为研究区位于西北干旱区,土壤贫瘠,盐碱化程度较高,导致叶片所获取的有效养分较少,这与罗艳等[20]和牛得草等[21]对塔里木河上游、阿拉善地区荒漠植物的研究结果一致。叶片C、N、P含量随着土壤含水率的降低而升高,受土壤水分限制,仅低水分梯度表现出了较高的叶养分浓度,说明叶片对干旱的响应策略不同[22]。高、中水分梯度受干旱影响程度相对较弱,能够凭借自身的渗透能力进行调节,而低水分梯度叶片在重度干旱条件下对干旱胁迫做出了响应。有研究表明干旱胁迫机制不仅会影响叶片C向淀粉转化,还能够使叶片中蛋白质和游离氨基酸升高维持细胞渗透平衡[23],所以在干旱条件下叶片会保持较高的C、N、P浓度,这也是植物在异质生境下的选择性适应策略。
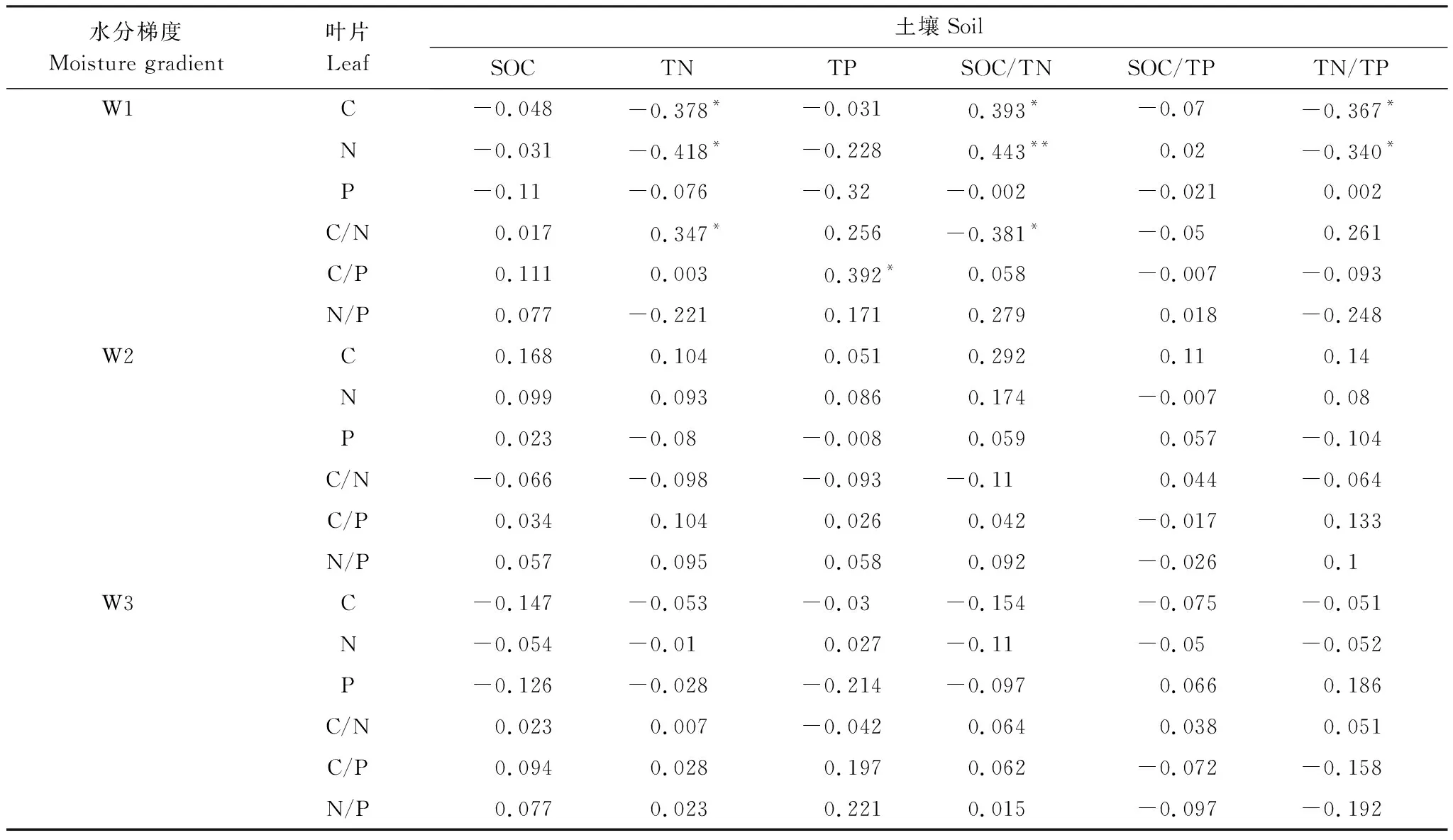
表2 不同水分梯度芦苇叶片和土壤C、N、P化学计量特征的相关性
叶片C/N、C/P是衡量植物养分利用效率的重要指标[24]。叶片平均C/N为21.27,变化范围在20.84—21.69,接近全球(22.5)[7]和黄土高原植物平均水平(21.2)[25]。叶片平均C/P为415.68,变化范围为344—455,远高于全球和黄土高原平均水平(232和312)[7,25]。该结果与Aerts和Chapin III[26]的研究结果一致,植物维持较高的叶片C/N、C/P有助于提高在干旱和营养不良环境中的竞争力。C作为结构性元素,其变异性较小,影响C/N、C/P值的主要因素是N和P[21],较高的C/N、C/P值代表植物N、P利用效率较高[27],相较于全球和黄土高原,本研究区P利用效率较高,这与研究区植物生长受限元素有较大的关系。Koerselman和Meuleman[8]认为可以用N/P阈值指示植物生长受N限制还是P限制,当N/P>16时,植物受P限制,当14 叶片C/N、C/P随着土壤含水率的升高而升高,说明高水分梯度芦苇在具有资源利用优势的条件下,与土壤达到了较高的平衡状态,叶片也具有较高的养分利用效率[13]。高水分梯度叶片C/P、N/P显著高于低水分梯度(P<0.05),而C/N不同梯度间无显著差异(P>0.05),这是因为植物中C/N相较于C/P、N/P具有更好的稳定性。Yang等[31]的研究表明,C与N之间的相互作用不依赖于生态系统的类型,因为N主要以有机N的形式存在于土壤和植物中。 土壤化学计量特征作为判断土壤有机质和质量的重要指标,可以指示和预测养分在土壤中的循环和变化趋势[6]。本研究土壤C、N、P平均含量(8.3、0.58 mg/g和0.42 mg/g)远低于全国平均水平(11.12、1.06 mg/g和0.65 mg/g)[20],可见研究区土壤总体养分含量较低。土壤中的营养元素尤其是C、N主要来源于土壤有机质和凋落物分解[20],芦苇作为研究区的单一优势种,生物量虽大,但因研究区干旱少雨,土壤盐渍化严重影响较难分解,凋落物反馈土壤受阻,影响土壤中有机质积累,因此土壤中C、N含量较低;P是一种沉积性元素,通常被认为是淡水湿地系统中最有限的养分,受成土条件和气候影响[2],研究区内的沙质土不利于P的积累,因此P含量较低。 土壤C、N和P随着土壤含水率的升高而升高,土壤含水率对土壤N的转化和有效性有积极的影响,并且土壤微生物在缺氧条件下活性较差,减缓了有机质的分解和氮的浸出,所以高水分梯度土壤C、N含量较高;系统的P输入主要依赖于生物过程、岩石风化和淋滤以及沉积矿物中P的赋存,相对较少依赖于土壤中的迁移[2],相比低水分梯度,高水分梯度土壤长期受水淹影响有利于P的储存,土壤P含量相对较高。 研究表明,土壤C/N与其有机质分解速度呈负相关[13],本研究中土壤C/N平均值(14.0)高于全球[32]及我国[20]土壤C/N平均水平(13.3和12.0),说明研究区土壤有机质分解速率相对缓慢。不同水分梯度土壤C/N表现为高水分梯度<中水分梯度<低水分梯度,且高水分梯度土壤C/N显著低于中、低水分梯度(P<0.05),反映了不同梯度内土壤有机质分解速度有差异,中、低水分梯度有机质分解速度较慢,主要是因为中、低水分梯度高盐碱、低水分的土壤环境抑制土壤中微生物群落活性(如表1),减缓了有机物的矿化分解[33]。土壤C和N随土壤含水率的升高而升高,而C/N呈相反趋势,可以看出研究区土壤中C和N的积累并不是同步的。此外,有机质分解特征与土壤C/N变化趋势相反,与N/P变化一致,这与王维奇等[34]对闽江河口湿地的研究结果一致。土壤C/P可以用来表示P有效性高低[13],不同水分梯度土壤C/P表现为中水分梯度>高水分梯度>低水分梯度,在一定程度上中水分梯度土壤P有效性可能要高于高、低水分梯度。土壤平均N/P(1.3)低于全球[32]及全国[20]平均水平(5.6和2.15),这表明N对土壤的制约作用要大一些,因为相对于P植物更倾向于保留N,特别是当土壤的N/P较低时[35]。Drenovsky和Richards[36]认为植物生长受到特定元素的限制原因是植物对特定元素的吸收率较低,而不是土壤缺乏该元素。植物与土壤作为生态系统的两大组成部分,它们的限制元素存在着较大的差异,这可能与它们不同的氮磷循环机制有关[2],这也是研究区叶片与土壤限制元素不同的原因。土壤N/P可以作为养分制约类型的有效预测指标[35],高水分梯度土壤N/P显著高于低水分梯度(P<0.05),说明本研究区内不同水分梯度土壤之间可能产生了不同养分限制类型,一个限制性影响因素可能会改变整个生态系统的功能[37],因此有必要进行进一步研究(如氮磷添加的控制实验[38])来确定土壤或植物中的限制性因素。 植物生态化学计量特征更倾向于是一种物种性状,与物种自身以及对环境的适应性有关[20]。在低水分梯度,叶片N、C/N和土壤N、C/N表现为显著负相关(P<0.05),这是因为土壤含水率较低时,土壤表层泥炭、草木残体及凋落物易氧化分解[39],导致大量养分损失,土壤亏损元素得不到补给而表现为负相关。叶片C、P、C/P、N/P和土壤C、P、C/P、N/P无显著相关性,叶片C主要来源于大气,而与土壤C无显著相关性;有研究表明,植物叶片P含量和土壤P含量密切相关,土壤中P浓度的增加往往导致植物P浓度的增加[2],这与本研究中结果不一致,这可能是低水分梯度土壤P有效性较低,叶片对P的利用率较低。低水分梯度叶片C、N、P养分浓度高于高、中水分梯度,而对土壤中C、N、P养分利用率较低,一方面可能与植物对养分的吸收利用及再分配有关,另一方面可能是低水分梯度土壤含水率较低,植物与土壤间的养分循环较弱而引起的[14]。 本研究中仅低水分梯度叶片N、C/N和土壤N、C/N显著负相关,高、中水分梯度叶片与土壤C、N、P化学计量特征相关性均不显著(P>0.05),这与塔里木河上游地区[20]及古尔班通古特沙漠地区[40]的结果一致,与呼伦贝尔沙地、毛乌素沙地[41]和黄土丘陵沟壑区[30]的结果不一致,说明在干旱或半干旱地区植物与土壤养分循环不仅受土壤水分影响,还受地形因子和植物种类的影响。崇明湖滩涂湿地[42]植物叶片与土壤C、N、P化学计量特征有显著相关性,而松嫩平原[43]及云南干热河谷地区[24]植物叶片与土壤C、N、P化学计量特征相关性不显著,这说明干旱地区和湿润地区叶片与土壤C、N、P化学计量特征相关性随生态系统类型和研究尺度的不同而不同。 植物体的营养元素主要来源于土壤,但是土壤中的养分却不一定是影响植物养分含量的主要因素。本研究选取对环境变化敏感的叶片来反映植物与土壤的密切关系,对于凋落物及土壤特性的研究较少,而凋落物是植物反馈土壤的重要环节,因此有必要结合凋落物及土壤特性进一步研究不同水分梯度植物与土壤的生态化学计量特征。 通过对不同水分梯度芦苇叶片与土壤C、N、P生态化学计量特征及其关系对比分析,发现研究区芦苇叶片和土壤养分整体偏低,土壤受N元素制约,而植物受P元素限制;与低水分梯度相比,高、中水分梯度土壤有效养分较高,植物与土壤达到较高的平衡状态,受土壤影响较小;而低水分梯度叶片受干旱胁迫和土壤养分制约,与土壤相关关系较为显著,并且能够保持较高的养分含量,体现了干旱区湿地植物独特的适应机制。3.2 不同水分梯度土壤C、N、P生态化学计量特征
3.3 不同水分梯度芦苇叶片与土壤C、N、P生态化学计量学的相关性
4 结论
