西南丘陵地区紫色土酸性对无患子幼树生长和光合特性的影响
2020-07-17田秋玲乐佳兴吴焦焦
田秋玲,乐佳兴,吴焦焦,张 文,高 岚,钱 春,刘 芸,*
1 西南大学资源环境学院, 重庆 400700 2 西南大学园艺园林学院, 重庆 400700
无患子(SapindusmukorossiGaertn.)是无患子科(Sapindaceae)无患子属(Sapindus)落叶乔木,其种仁含油率高达40%,可用于提炼生物柴油和制备高级润滑油,果皮提取的皂苷具有洗涤降污、洗脱重金属、抗菌抗肿瘤等多种生物活性[1],材质致密、花纹美观,富含的天然皂素可自然防腐防虫,是木质家具和工艺品的优良原料[2],也因树形优美、树干通直、秋季叶色金黄被广泛用作行道树[3]。可见,无患子是集医药化工、生物质能源、工业制造和园林绿化于一体的多用途经济树种。无患子属在全世界共13种,我国有4种1变种,适宜热带、亚热带地区生长,分布中心位于热带美洲,我国淮河流域以南是边缘分布区,主要分布于东部、南部、西南部,为低山、丘陵及石灰岩山地常见树种[4- 6]。近年来,无患子在重庆、贵州、云南等西南贫困山区的退耕还林、植被恢复中作为重要经济生态树种被大力推广种植。
西南丘陵地区退耕还林土地以紫色土居多,紫色土是由紫色母岩发育而成的幼年土,风化程度低,原生矿物和盐基离子含量丰富,粘土矿物以蒙脱石和水云母为主使得土壤钾含量丰富,种植经济林木效益高[7- 8]。但有研究表明,近年来紫色土受酸沉降和氮肥施用影响,已出现不同程度酸化,紫色土强酸化(pH<4.5)后具有交换性酸和交换性盐基离子同时增多的“双面性”酸化特征,交换性酸含量远多于同等酸度的黄壤和砖红壤[9- 11]。土壤是植物完成生活史最重要的介质,中性紫色土尚可维持较高肥力水平,但酸性紫色土由于交换性酸含量极高并且活化了重金属离子而对植物根系产生毒害[12- 13],不利于植物生长发育。目前,有关无患子的研究多集中于化学成分的作用分析和提取工艺[14- 15]、种质资源调查[5]、物候期观测[16]等方面,而关于原料林培育中受酸性土壤胁迫则未见研究报道。
光合作用是绿色植物将光能和化学能转化为有机物的唯一途径,植物受到逆境胁迫时,光合作用减弱,生产力低,植物生长发育受阻。因此,本试验研究了重庆市紫色土不同程度酸性区(中性区为对照)3年生无患子幼树的生长、光合特性差异,初步探讨土壤酸性对无患子生长及光合作用的影响,为无患子在西南地区紫色土推广种植之适生区选择提供理论依据。
1 材料与方法
1.1 紫色土酸性区试验选址
根据李忠意等[17]研究,重庆市紫色土土壤pH范围为3.7—8.8,土壤 pH在6.5—7.5之间属中性,5.5—6.5范围内以交换性H+为主组成的交换性酸含量极低,属弱酸性,土壤pH < 5.5时交换性酸和交换性Al3+随土壤pH降低呈指数增长且交换性Al3+占土壤交换性酸的比重不断增加[18],属强酸性。本试验根据前人研究,结合前期调查结果,分别选址重庆市北碚区西南大学水保园中性紫色土区(简称S1)、垫江县长龙镇长堰村紫色土弱酸性区(简称S2)、合川区西山坪镇野猫坪村紫色土强酸性区(简称S3),上述3个试验区均属于川东平行岭谷区,丘陵地形,亚热带湿润季风气候,年均降水量1230 mm,集中在6—8月,年均日照时间1245.5 h,最高温40℃,最低温4℃,年均温17℃。基本信息见表1。

表1 试验区概况
1.2 实验设计
3个试验区均采用完全随机区组设计,在每个试验区选择地势平坦的地方设置3个实验小区,小区规格12 m × 12 m,每小区内重复16株,株行距3 m × 3 m。于2016年11月选取长势一致的1年生健壮幼苗,平均株高和地径分别为(95.71±3.07) cm、(10.75±0.18) mm,按试验设计随机定植于小区,共144株。后期采用相同田间管理措施,均无任何形式的肥料添加。2018年7月初,同时对3个试验区的无患子进行生长和光合参数测定。
1.3 土壤理化性质测定
分别于2016年11月(幼苗种植前)、2018年7月采集试验区土壤,每小区以“S”型采样法采集根际耕层(0—20 cm)土壤混匀后用四分法保留约1 kg土壤样品,带回实验室自然风干后过2.00、1.00 mm和0.25 mm尼龙筛备用。采用杨剑虹等[19]的方法测定土壤理化性质。
1.4 生长、叶性状指标测定
每小区随机选取10株,用卷尺(1 cm精度)测量株高和冠幅,游标卡尺(精度0.02 mm)测量地径(从树基部向上1 cm处树干直径)。利用智能叶面积测量系统(浙江托普云农科技股份有限公司,中国)测量叶片的叶面积(LA),扫描结束后置于65 ℃烘箱内烘至恒重,用电子天平(0.0001 g精度)称量叶干质量,比叶面积(SLA)= 叶面积/叶干重,每小区重复5株,每株20片,结果取平均值。本实验中测量叶片均为树冠中上段向阳方向复叶顶端的第3或第4片健康小叶。
1.5 叶片全氮(TN)、全磷(TP)、全钾(TK)含量测定
取小叶装入冷藏箱带回实验室,65 ℃烘箱内烘干至恒重,磨细过2 mm尼龙筛备用,每小区重复3株,每株200片。叶片TN采用凯氏法测定,叶片TP用H2SO4-HClO4消化后,钥兰比色法测定,叶片TK采用火焰分光光度计测定[19]。
1.6 光响应曲线及气体交换参数测定
采用Li- 6800光合测定系统(Li-COR,美国) 2 cm × 3 cm叶室测定叶片光响应曲线,通过LED红蓝光源设置12个光合有效辐射(PAR,μmol m-2s-1)强度梯度:1800、1500、1200、1000、800、600、400、200、150、100、50和0 μmol m-2s-1,CO2浓度400 μmol m-2s-1(由小型 CO2钢瓶提供),温度25℃,相对湿度60% ± 5%,气体流速500 μmol/s,测定前在1200 μmol m-2s-1光强下诱导15 min,每个光强下稳定2 min后记录数据。利用叶子飘[20]的光合计算方法,使用直角双曲线修正模型拟合并计算最大净光合速率(Pnmax)、光补偿点(LCP)、光饱和点(LSP)、暗呼吸速率(Rd)、表观量子效率(AQY) 及拟合决定系数(R2)。每小区重复3株,每株测量3片,结果取平均值,同时标记测量叶片。
2018年7月初选取连续晴天9: 00—11: 30,利用上述Li- 6800光合测定系统测定气体交换参数,空气流速为500 μmol/s,每个点稳定2 min后读数,记录净光合速率(Pn) 、蒸腾速率(Tr) 、气孔导度(Gs) 、胞间CO2浓度(Ci),叶室CO2浓度(Ca),气孔限制值(Ls)= 1 -Ci/Ca。测定的叶片和重复数与上述光响应曲线相同。
1.7 光合色素含量及叶绿素荧光参数测定
利用JUNIOR-PAM便携式脉冲调制荧光仪(WALZ,德国)测定叶片荧光特性,测定前暗适应20 min,后照射检测光测得初始荧光(Fo);然后用强饱和脉冲光(10000 μmol m-2s-1) 激发,使原初电子受体QA全部处于还原状态,测定最大荧光(Fm),可变荧光Fv=Fm-Fo,最大光化学效率(Fv/Fm) 、光化学猝灭系数(qP) 、非光化学猝灭系数(qN) 、电子传递速率(ETR)和实际原初光能捕获效率(Y(Ⅱ))均由仪器计算给出。测定的叶片和重复数与光响应曲线相同。
叶绿素荧光测定完成后,立即摘下叶片,-80℃液氮低温保存,带回实验室立即测定光合色素,总叶绿素(Chls)、叶绿素a(Chl a)、叶绿素b(Chl b)、类胡萝卜素(Cars)含量采用乙醇提取法测定[21]。
1.8 统计分析
使用Excel软件完成数据整理、制表和绘图,SPSS 23.0统计软件进行单因素(one-way ANOVA)方差分析、Pearson相关性分析和Duncan多重比较。
2 结果与分析
2.1 土壤酸性对紫色土速效养分和叶片氮(TN)、磷(TP)、钾(TK)含量的影响
由表1、2、3知,各试验区土壤TN和SOM含量为S3 > S1 > S2,与土壤pH均无显著相关性,酸性区土壤TP、TK含量低于中性区,与土壤pH极显著正相关(P<0.01)。与中性区相比,酸性区土壤AN、AP含量极显著增加(P<0.01),且均与土壤pH极显著负相关,酸性区土壤AK含量极显著减少,与土壤pH极显著正相关,不同试验区之间差异极显著(P<0.01)。与中性区相比,酸性区叶片TN和TK含量显著减少,土壤越酸,含量越低,叶片TP含量在弱酸性区最高,不同试验区之间叶片TN、TP和TK含量差异均极显著(P<0.01)。
2.2 土壤酸性对无患子幼树生长及叶性状的影响
如表4所示,株高为S1 > S2 > S3,不同试验区之间差异极显著(P<0.01),地径在S2最大,S3最小,S1与S2差异不显著(P>0.05),酸性区的幼树冠幅显著高于中性区(P<0.05)。各试验区SLA、LA均是:S2 > S1 > S3,LA在不同试验区之间差异极显著(P<0.01)。
2.3 土壤酸性对无患子幼树叶片光合色素含量及叶绿素荧光参数的影响
由表5可知,中性区幼树叶片Chl a、Chls、Cars含量以及Chl a/b比值最大,而Chl b含量和Chls/Cars比值最小,且与酸性区差异极显著(P<0.01),酸性区之间无显著差异(P> 0.05)。与中性区相比,酸性区的 Y(Ⅱ)、ETR和qP显著增大(P<0.05),其中Y(Ⅱ)和ETR在酸性区之间无显著差异(P>0.05),不同试验区qP差异极显著(P<0.01)。酸性区Fv/Fm和qN显著小于中性区,不同试验区间Fv/Fm差异极显著(P<0.01),qN表现为中性区极显著高于酸性区(P<0.01)。

表2 不同试验区土壤速效养分和叶片氮(TN)、磷(TP)、钾(TK)含量

表3 土壤养分与土壤pH相关性分析
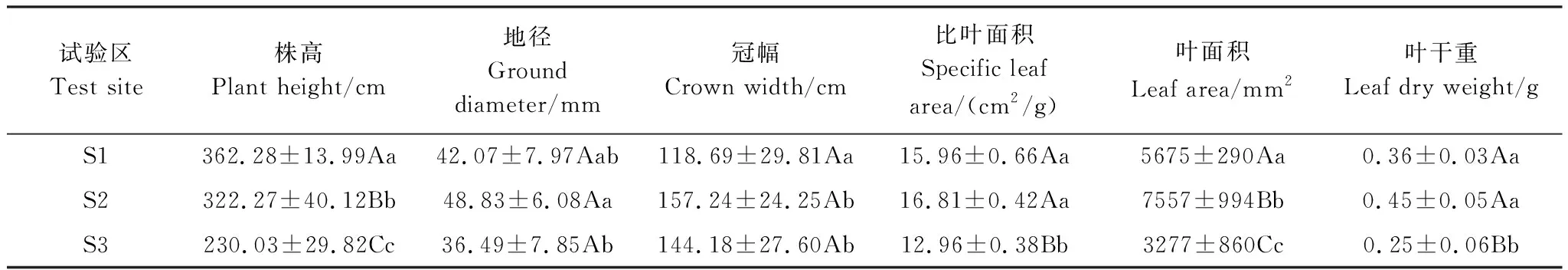
表4 不同试验区无患子幼树生长及叶性状指标
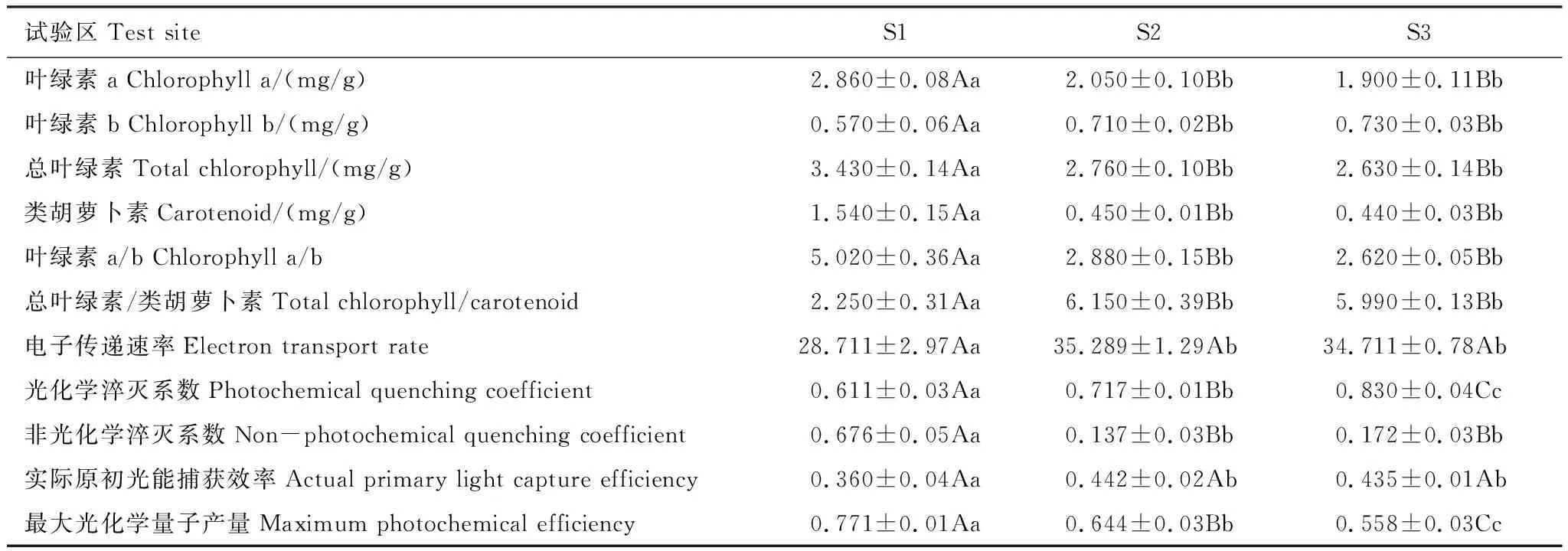
表5 不同试验区无患子幼树叶片光合色素含量及叶绿素荧光参数
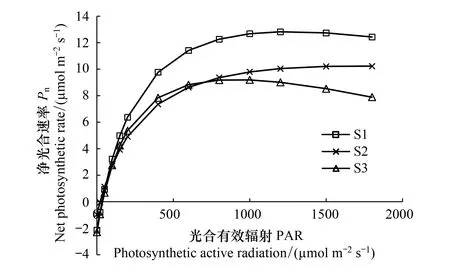
图1 不同试验区无患子叶片净光合速率Pn对光照强度的响应Fig.1 Response of net photosynthetic rate Pn to light intensity in of leaves of S. mukorossi in different test areas
2.4 土壤酸性对无患子幼树叶片光响应曲线及气体交换参数的影响
在CO2恒定400 μmol/mol浓度和0—1800 μmol m-2s-1光强范围内测得光响应曲线并得出拟合参数。由图1可知,不同试验区叶片Pn随PAR的变化趋势基本一致,PAR从0增至400 μmol m-2s-1时,叶片Pn快速增加,PAR>400 μmol m-2s-1后,Pn缓慢增加,S3在PAR趋近1000 μmol m-2s-1时Pn出现缓慢下降趋势。
如表6所示,与中性区相比,酸性区Pnmax显著减小,与中性区差异极显著(P<0.01),S3的LSP极显著下降(P<0.01),各试验区AQY、LCP和Rd均为S3 > S1 > S2,其中 AQY和Rd在不同试验区之间差异极显著(P<0.01);酸性区Pn极显著下降(P<0.01),试验区之间差异达极显著水平(P<0.01),Tr表现为S2极显著高于S1和S3(P<0.01),S1和S3之间差异不显著(P>0.05),Ci和Gs均为S2 > S1 > S3,S1和S2极显著高于S3(P<0.01),Ls为S3 > S1 > S2,S1和S2极显著低于S3(P<0.01)。
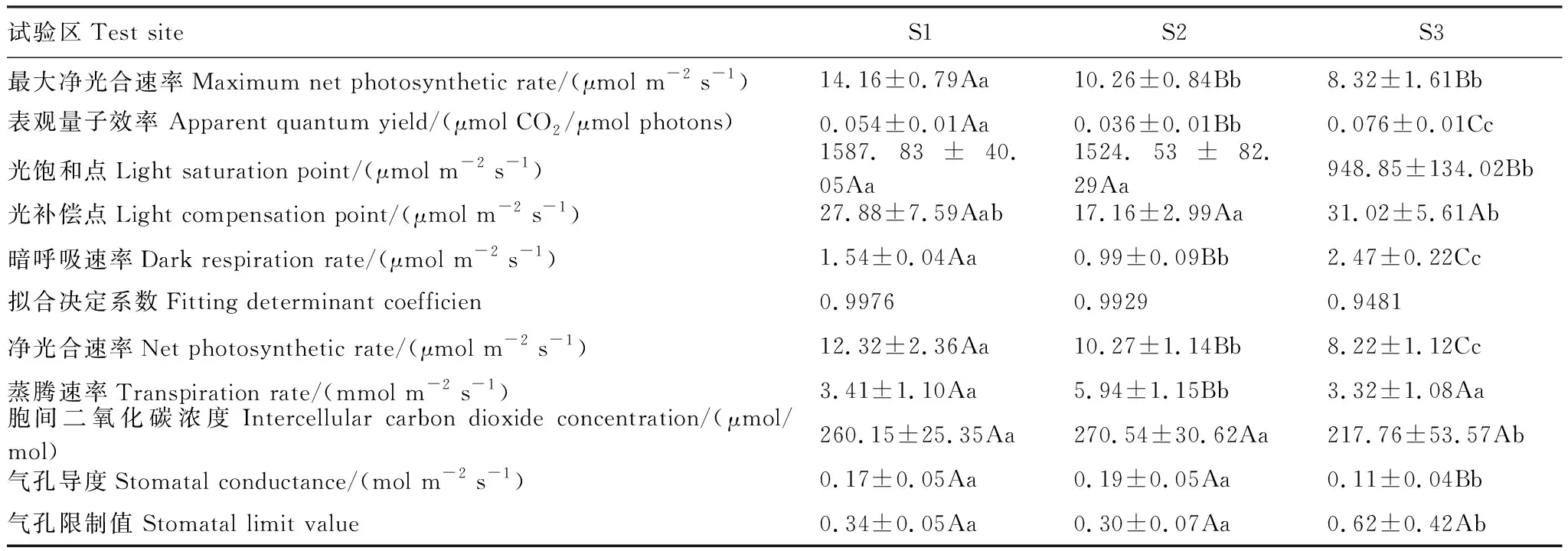
表6 不同试验区无患子幼树叶片光响应曲线及气体交换参数
3 讨论
土壤pH能够影响或指示土壤养分的转化、释放、有效性及植物根系吸收养分的能力。有研究表明,酸性紫色土中氮素更易矿化,土壤pH越低,氮矿化量越多[22- 23]。肖懿、Zhang等[24- 25]认为土壤酸性会导致固定的磷素溶解,与中性和石灰性紫色土相比,酸性紫色土磷素最大吸附量、吸附缓冲容量低,磷素解吸能力高,磷素释放风险大。本研究结果发现,紫色土酸性促进了土壤氮磷元素向其有效态转化,使得土壤AN、AP含量高。这与李忠意等[17]的研究结果一致。土壤酸性会加速土壤矿物分解,促进矿质态钾转变成有效态钾,而钾素极易被淋失,从而造成钾素养分库损失严重[26- 27]。本研究中,紫色土酸性区土壤AK与TK含量低可能与其长期酸性条件下钾素转化为有效态后大量淋失有关。
叶片氮作为构建叶绿素的主要成分,当其显著下降时必然导致叶绿素含量降低,不利于光能的吸收和转化[28]。本研究结果显示,中性区幼树叶片较高的TN含量促进了叶绿素合成,叶片高的Chls、Chl a含量和Chl a/b比值与其高光合能力相对应[29]。Chl a和Chl b分别表征叶片光能利用和捕获能力的指标[30],酸性区生长的无患子通过增加Chl b含量提高叶绿体弱光捕获能力,而Chl a含量极显著减少造成叶绿体光电转化效率降低,进而光合速率显著下降。Lichtenthaler等[31]认为陆地阳生植物叶片Chl a/b比值是3.0—3.4,Chls/Cars比值是3.8—4.4;阴生植物Chl a/b比值为2.4—2.7,Chls/Cars比值为4.8—5.7。本研究中,中性区叶片Chl a/b和Chls/Cars比值极具阳生植物特性,而酸性区叶片Chl a/b和Chls/Cars比值更趋向阴生植物特性。实际上,无患子属阳生植物,喜全光照。与中性区叶片色素含量相比,土壤酸性导致无患子色素含量失衡,对光照的需求表现出阴生植物特性,即更偏爱相对低的光照和散射光。可见,土壤酸性可通过影响无患子色素含量和比值,进而影响其光合能力。LSP和LCP的高低能够反映植物的需光特性和环境适应能力[32]。本研究表明,强酸性区LSP低,强光利用能力弱,LCP和Rd较高,属于低生产高消耗,生态适应性弱;弱酸性区LSP高,LCP和Rd最低,光能利用能力强,属于高生产低消耗,生态适应性强;中性区LSP高,LCP和Rd较高,弱光利用能力差,消耗较大,但Pnmax最大,叶片有机物质积累多,生态适应性强。可见,本实验中,紫色土中性区和弱酸性区是无患子生长适宜区。
叶绿素荧光参数可反映植物受逆境胁迫的光合机构响应方式[33]。Fv/Fm是表征PSII反应中心内禀光能转化能力的指标,非胁迫条件下一般为0.75—0.85[34]。本研究中,酸性区Fv/Fm极显著降低,表明无患子叶片光合潜能受土壤酸胁迫影响而下降,与张翠萍等[35]研究结果相同;qN是PSⅡ反应中心以热形式耗散过剩光能的比例[36],无患子幼树叶片应对土壤酸胁迫没有增加热耗散来消耗过剩激发能,与李清雪等[37]研究结果相似。此外,中性区叶片高的Cars含量对其通过叶黄素循环耗散过剩光能以应对夏季高光环境下光合机构免受破坏有积极作用[38- 39]。本实验范围内,酸性区qP、Y(II)和ETR均显著高于中性区,说明酸性区叶片吸收的光能用于光化学传递的份额大,酸性区无患子叶片在光能捕获量不足条件下通过调整光合结构,以提高光合电子传递速率和光能捕获效率,实现吸收的光能利用效率最大化。可见,无患子在土壤酸性胁迫下,具有一定的主动适应逆境能力。
叶性状与植物资源利用策略及生存环境适应能力间存在一定协调性。无患子在土壤酸胁迫影响下,其叶片表现出具有一定适应性的表型可塑性。钾在增大叶面积、延长叶的功能期、提高叶片叶绿素含量和气孔开张灵活度等方面起重要作用,从而提高净光合速率,促进有机物质积累[40- 41]。本研究中,强酸性区叶片TK含量低可能与其Tr、Gs下降有关,同时 LA、SLA、Chls和Chl a含量减小,使得光合生产力下降,植株生长受阻;弱酸性区幼树叶片大的LA、SLA有利于拦截光能[42],从而补偿较低光量子通量密度造成的光合速率下降,薄而大的叶片Tr和Gs高,使其运输水分、养分的量和速率增加,并且CO2供应充足,与其高光合速率相适应[43],有利于光合产物积累,表现出地径和冠幅均最大。酸性区幼树株高显著减小,但冠幅显著增大,可能是土壤酸性影响了植物激素合成而削弱了无患子树体的顶端优势,关于这一点,有待进一步研究。
4 结论
紫色土酸性通过影响土壤养分转化、根系吸收能力显著降低了无患子幼树的光合能力;强酸性区幼树叶片氮、钾含量低,导致其光能利用能力差,光能捕获总量低,有机物质消耗大,不利于无患子生长;弱酸性区幼树叶片通过增大光能捕获面积,提高PSⅡ光合电子传递速率和光能捕获效率,降低有机物质消耗,表现出较强的生态适应性。研究结果表明,无患子在重庆紫色土区种植的pH适宜范围为5.5—7.5,而在pH值低于5.5的紫色土种植无患子,对其生长发育不利。
