桢楠幼树光合特性对镉胁迫的响应
2020-07-17铁得祥胡红玲喻秀艳舒奕嘉
铁得祥,胡红玲,*,喻秀艳,2,舒奕嘉,张 健
1 四川农业大学林学院, 成都 611130 2 自贡市林业重点工程管理中心,自贡 643000
植物的光合作用对重金属较为敏感[1],对于非耐镉植物而言,光合合色素、气体交换参数和叶绿素荧光参数等光合特性指标在Cd胁迫下均会呈现出显著降低趋势[2],其原因主要有以下几方面:Cd会阻碍叶绿素的生物合成[3]、破坏光合器官的结构和功能,显著降低植物的光合作用速率;还能干扰电子转移和ATP的合成过程、抑制暗反应阶段的关键酶活性[4- 5];此外,也能通过改变植物叶肉细胞的水势和膨压,来控制植物的气孔行为[6],进而降低植物的气体交换速率,使得部分气体交换参数值下降。但对与耐Cd植物而言,较低浓度镉胁迫下,植物的光合作用并不会受到显著抑制,如宝山堇菜(Violabaoshanensis),龙葵(Solanumnigrum)等植物[7- 8];由此可见,Cd对植物光合作用的影响程度大小与植物对Cd的耐受程度密切相关。此外,不同耐Cd植物对镉胁迫的光合特性响应也存在明显差异,如龙葵和印度芥菜 (Brassicajuncea)的叶绿素含量和净光合速率在较低Cd浓度胁迫与正常生长环境下并无显著差异,但部分气体交换参数值(蒸腾速率和气孔导度)却呈现出相反的变化趋势[7,9];再如球果蔊菜(Rorippaglobosa)的叶绿素含量随Cd胁迫程度的加重呈先升后降趋势[10],而龙葵的叶绿素含量则呈逐渐下降的趋势[7]。综上表明,不同植物的光合特性对Cd胁迫的响应存在很大差异。
桢楠(Phoebezhennan)属樟科(Lauraceae)楠属(Phoebe)植物,是我国特有的珍稀树种,在我国主要分布于四川盆地、贵州北部及湖北地区。该树种多为高大的常绿乔木,生命周期长、幼龄期生长快,适应性较强,目前在绿化工程中广泛应用[11]。也因其特殊的材质、纹理特征和医药成分,在林业、轻工业、医药等行业都具有较高的经济价值[12]。目前对于桢楠光合特性的研究多集中在干旱、施肥、水淹胁迫等方面[13- 14],但都未从叶绿素荧光层面去进行更加深入的探究,作为植物光合作用与环境关系的内在探针[15],通过分析桢楠幼树体内叶绿素荧光参数和光合作用参数的相互关系,能更加全面地了解桢楠幼树的光合作用机理,如植物叶片的电子传递、转移过程,对吸收光能的利用情况等[16- 17]。目前由于工业排放和农药化肥的大量施用,重金属污染风险逐步加重,尤其是在长江中下游地区Cd污染日趋严重,因此探讨桢楠幼树光合生理和叶绿素荧光对不同浓度Cd胁迫的响应情况,有助于更加全面的了解桢楠幼树对Cd的基本适应能力,为深入研究桢楠抗Cd胁迫机制提供理论依据,也可为选择适宜的桢楠栽培立地提供参考。
1 材料与方法
1.1 试验地概况
试验地设在四川农业大学(成都校区)教学科研基地,地理位置为东经103°51′29″,北纬30°42′18″,属亚热带湿润气候区,四季分明,气候温和,雨量充沛。年平均气温16.8 ℃,月平均最高温为25.9 ℃(7月),月平均最低温为6.9 ℃(12月),极端最高气温38 ℃(7月),极端最低气温-2 ℃(1月);累年平均相对湿度达84%;全年平均降水量为759.1—1155.0 mm;年平均总日照时数为840.2 h。
1.2 试验材料
1.2.1栽植土壤与容器
试验用土为农田沙壤土,其理化性质为:pH6.97、有机质27.51 g/kg、全氮2.31 g/kg、全钾3.88 g/kg、汞0.01 mg/kg、砷9.91 mg/kg、镉0.19 mg/kg、铅3.34 mg/kg、铬100.75 mg/kg、铜40.27 mg/kg。栽植容器均采用规格一致的塑料盆(盆口直径30 cm,底径20 cm,高23 cm),每盆等量装土8.5 kg(干土质量)。
1.2.2供试植株
为同一种源的2.5 a生桢楠幼树,无病虫害,平均株高118.55±8.80 cm,平均地径13.37 ±0.20 cm,每盆1株,移栽后常规水费管理,进行为期1 a的适应性培养。期间采用HH2土壤水分测定仪(ML2x,GBR)监测土壤水分状况,使土壤体积含水量保持在20%左右(即本试验用土田间持水量的80%)。试验开始前15 d,将所有待处理植株移入通风秀气的大棚内,并停止施肥,以防肥料对镉效应的表达产生干扰。
1.3 试验设计
采用单因素试验设计,根据四川省土壤环境调查数据[18],四川有较多地区土壤Cd含量达4.4 mg/kg,且少数污染严重地段镉含量高达22.20 mg/kg,加之近年来西南地区土壤Cd含量增幅较大[19],因此根据四川地区镉污染现状并结合未来可能的发展趋势,本试验共设置6个Cd处理水平:CK(0 mg/kg)、T1(10 mg/kg)、T2(20 mg/kg)、T3(30 mg/kg)、T4(40 mg/kg)、T5(50 mg/kg),其中Cd以CdCl2的形式水溶施入,用量见表1。为避免苗木移入塑料盆后的适应培育过程中一些植株因外界干扰生长受阻或意外死亡影响样本选择,每个处理重复9次,共计54盆。所有处理植株均放置在通风透气大棚内培养。试验于2017年6月初开始,以CdCl2与相同体积去离子水配成母液,然后稀释成不同处理浓度。为使镉施入方式与现实中镉排放过程相似,本次试验将各处理CdCl2等分为5份,分别于2017年7、9、11月,2018年1、3月施入。施入时,将配制的溶液均匀浇灌在盆内土壤表面,另用托盘垫于盆底收集可能渗出的CdCl2溶液,并将其浇回土壤中,对照浇等量的去离子水。于2018年5月初试验结束时,每个处理抽取3株重复,在每株新梢上相同叶位选取3片成熟功能叶,采用Li- 6800便携式光合测定仪(Li-Cor Inc.,USA)在不同时间分别测定各处理植株叶片的气体交换参数、光响应特征曲线以及叶绿素荧光参数,之后采集功能叶,装入密封袋,标记好之后置于冰盒中,即刻带回实验室进行叶绿素含量测定。

表1 重金属Cd处理水平
1.4 测定指标及测定方法
1.4.1光合色素含量测定
采用分光光度法(80%丙酮与无水乙醇1∶1混合提取法)测定[20]。
1.4.2光合气体交换参数测定
净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)使用Li- 6800便携式光合测定仪(Li-Cor Inc.USA)于2018年5月一个晴天上午9:00—11:00进行测定,设定参比室CO2浓度为400 μmol/mol,叶室温度为30 ℃,叶室相对湿度为50%,光照强度设定为环境光强(1000 μmol m-2s-1)。叶片均选用同方位成熟的功能叶,每个处理重复测定3株,每株测定3个叶片。
1.4.3光合-光响应曲线测定
使用Li- 6800进行测定,测定时间为2018年5月多个晴天上午(8:30—11:30),叶片选择标准与数量同上。光强梯度设定为1800、1500、1200、1000、800、600、400、200、150、100、75、50、25、0 μmol m-2s-1,叶室其他参数设定与测定气体交换参数的设定一致,通过预试验测定桢楠幼树的光饱和点在800 μmol m-2s-1左右,故首先将桢楠幼树叶片在800 μmol m-2s-1的光强下进行光诱导直至Pn稳定(大约5 min),然后使用光响应曲线自动测定程序进行测定。
1.4.4荧光参数测定
使用Li- 6800进行测定,叶片选择标准与数量同上,叶室参数设定与测定气体交换参数的设定一致。测量前将供试植株移入无光实验室进行暗适应,并用锡箔纸包裹待测叶片(标记测定叶片,并充分保证暗适应),待暗适应一整夜后,在第二天8:00开始荧光参数的测定,首先用一束小于0.1μmol m-2s-1的测量光(频率为50 Hz)照射,得到初始荧光F0;后打开一个饱和脉冲光(8000 μmol m-2s-1,100 Hz,1000 ms),得到暗适应下的最大荧光Fm,然后根据环境光强设定活化光,待F稳定后,此时的F即为稳态荧光Fs;然后再打开一个同样强度的饱和脉冲光,得到Fm′;同样打开远红光(25 μmol m-2s-1)得到F0′。根据测定数据,可计算得出PSⅡ潜在活性(Fv/F0)、PSⅡ最大光化学量子产量(Fv/Fm)、PSⅡ有效光化学量子产量(Fv′/Fm′)、PSⅡ实际光化学量子产量(ΦPSII)、电子传递速率(ETR)、光化学淬灭系数(qP)和非光化学淬灭系数(NPQ)[21]。
1.4.5PSⅡ吸收光能的分配比例
根据Demmig|Adams等[22]提出的方法计算:
(1)光化学耗散(P)=qP×Fv′/Fm′×100%;
(2)天线热耗散(D)=(1-Fv′/Fm′)×100%;
(3)过剩激发能(Ex)=(1-qP)×Fv′/Fm′×100%。
1.5 数据处理
使用Excel 2016软件(Microsoft Office, USA)进行数据处理,采用SPSS 22.0对数据进行单因素方差分析(One-way ANOVA),显著水平α设定为0.05,用Duncan′s法进行多重比较,最后用Sigmaplot 12.5(Systat Software Inc., USA)做图。
光合-光响应曲线测定结果采用双曲线修正模型拟合,同时获得最大净光合(Pmax)、光补偿点(LCP)、光饱和点(LSP)和暗呼吸速率(Rd)等光合特征参数[23- 24]。
2 结果与分析
2.1 镉胁迫对桢楠幼树叶绿素含量的影响
如图1所示,随着Cd胁迫程度的加重,叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素(Car)和叶绿素总量(Chla+b)均呈下降趋势。其中Chla、Chlb、Chl(a+b)分别在T3、T2、T3时显著下降(P<0.05),相比CK降幅分别达45%、51%、46%,Car含量在T1时显著下降,降幅达34%。表明低浓度(T1—T2)胁迫下,Cd对Chla、Chlb、Chl(a+b)含量无明显影响,但对Car影响显著;而高浓度(T3—T4)Cd胁迫显著影响了各类叶绿素含量。由图1还可知,Chla/b随Cd胁迫的增大逐渐上升,可见Cd对Chla的抑制程度比Chlb更弱一些。
2.2 镉胁迫对桢楠幼树气体交换参数的影响
如表2所示,随着Cd胁迫程度的加重,桢楠幼树的Pn、Gs、Tr均呈下降趋势,且分别在T1、T1、T2时显著下降了24%、31%、21%。且随Cd胁迫程度的加重而增加,Ci呈在增加趋势,LS呈降低趋势,并均在T4处理时显著高于(低于)CK,表明此时桢楠幼树叶片的固C能力减弱,且桢楠幼树叶片光合速率的降低主要由非气孔因素决定。
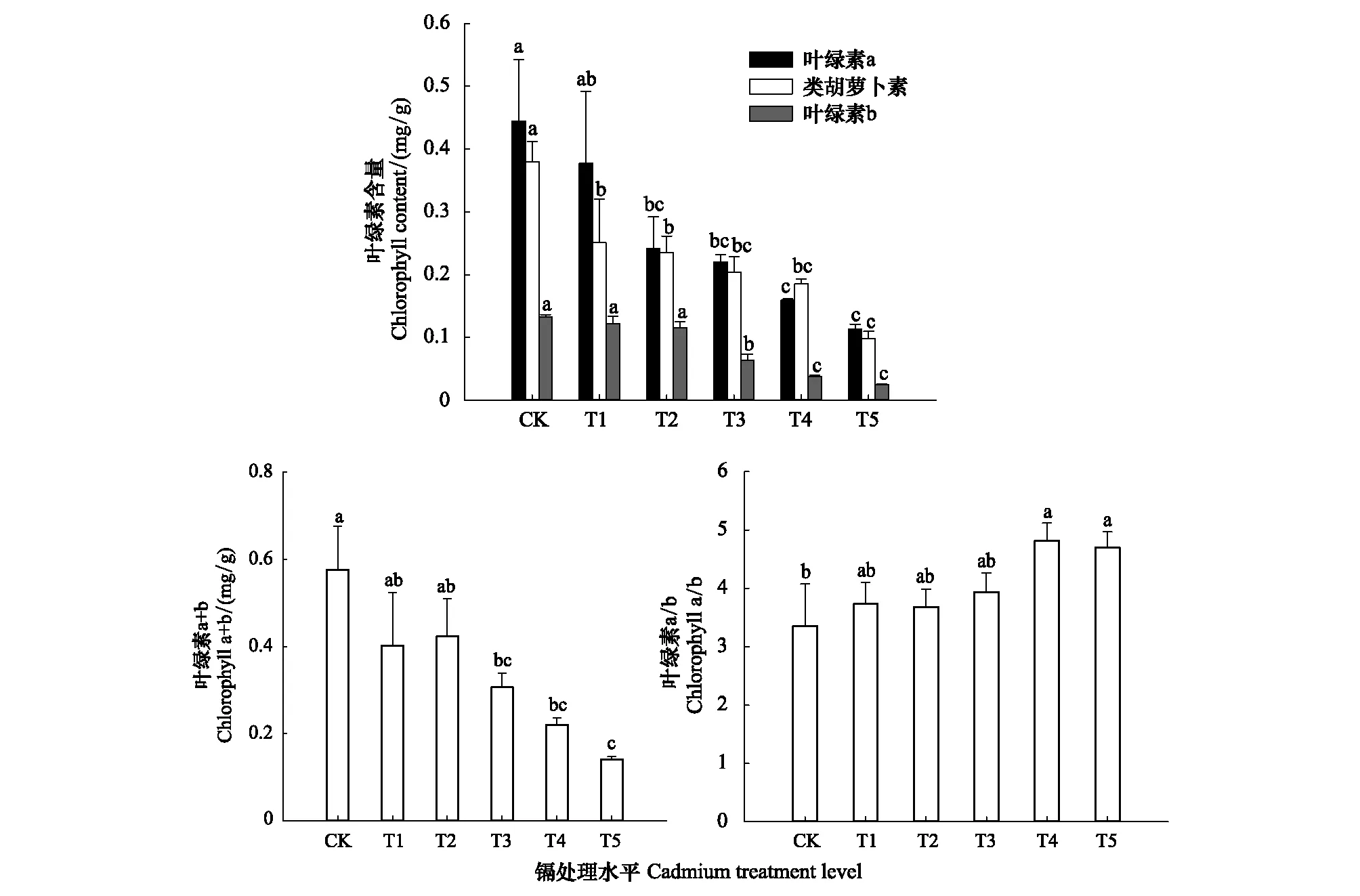
图1 不同水平镉处理下桢楠幼树叶片中色素含量变化Fig.1 Changes of pigment content in leaves of P. zhennan leaves under different levels of cadmium treatment图条上方不同小写字母分别表示镉处理间差异显著(P < 0. 05)
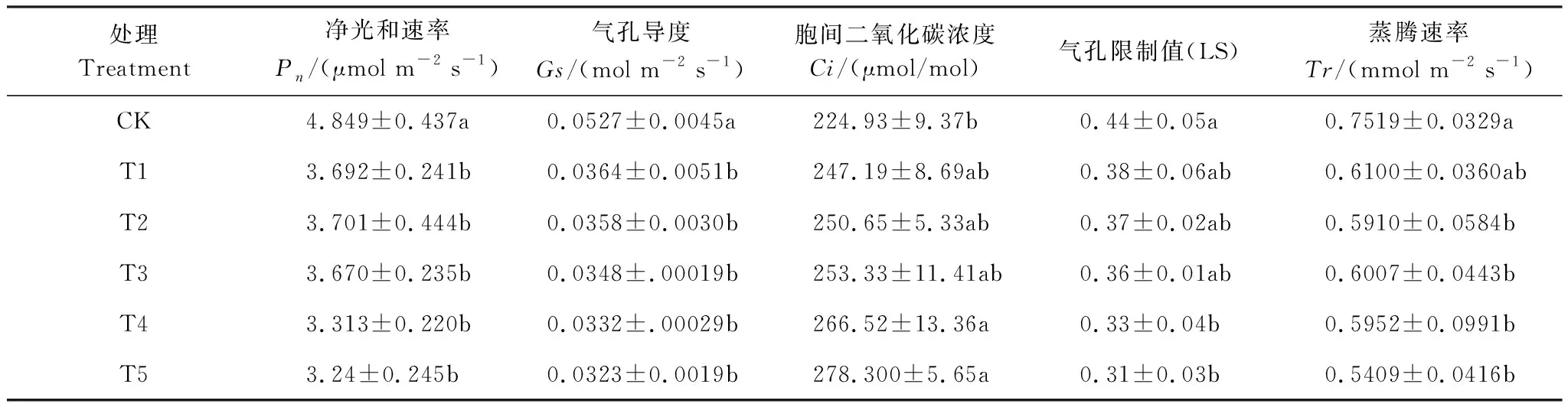
表2 不同水平Cd处理下桢楠幼树气体交换参数变化
2.3 镉胁迫对桢楠幼树叶片光合-光响应特征的影响
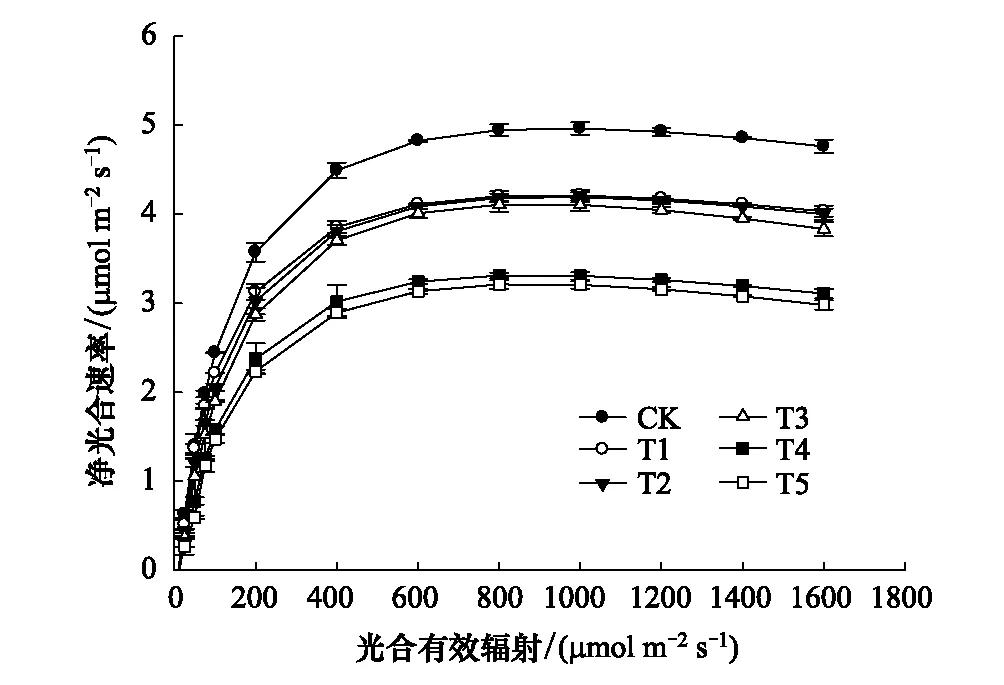
图2 不同水平镉处理下桢楠幼树叶片的光合-光相响应曲线 Fig.2 Photosynthetic-optical response curves of leaves of P. zhennan var.
由图2可知,在同一PAR下,随着Cd胁迫程度的加重,桢楠幼树对光能的利用率逐渐降低,最大净光合速率(Pmax)呈下降趋势。在同一镉处理下,当PAR≤100 μmol m-2s-1时,Pn均随光合有效辐射的增强而急剧增大,二者基本呈直线关系,此时PAR是限制桢楠幼树光合作用的唯一因素;随着PAR的持续增强,各处理植株Pn仍呈上升趋势;当PAR≥800 μmol m-2s-1时,Pn基本趋于平缓,此时CO2浓度成为限制桢楠幼树光合作用的唯一因素,且CK的最大Pn均显著高于Cd处理植株(表7),T2与T3最大Pn也显著高于T4与T5,而T2与T3,T4与T5间则无明显差异。由表7可知,随着Cd胁迫程度的加强,桢楠幼树的表观量子效率(AQY)逐渐降低,光补偿点(LCP)逐渐升高,表明桢楠幼树光合机构遭受一定程度的破坏,其对弱光的利用能力、干物质积累能力等均减弱。
2.4 镉胁迫对桢楠幼树叶绿素荧光参数的影响
如表4所示,Fv/Fm、Fv/F0在T1—T4处理范围内无显著变化,在T5处理时显著降低;ΦPSⅡ、ETR、qP均随Cd胁迫程度的加重呈下降趋势,T1—T2时显著降低;NPQ呈升高趋势,并至T4处理时显著高于CK。由此可知,在T1—T4处理时,桢楠幼树叶片仍能保持正常的光能转化效率及稳定的PSⅡ潜在光活性,但叶片PSⅡ光反应中心对Cd较为敏感,Cd胁迫下其受损明显,开放程度降低,但桢楠幼树也会尽力提高PSⅡ反应中心的热耗散能力(NPQ上升),以此减轻光抑制所产生的伤害。

表3 不同水平镉处理下桢楠幼树叶片光响应曲线特征参数
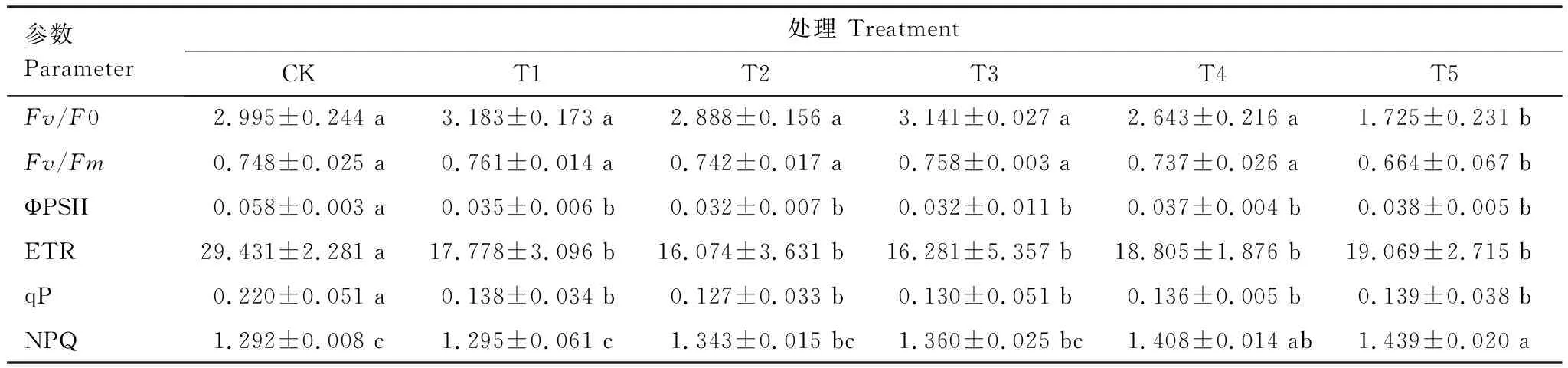
表4 不同水平镉处理下桢楠幼树荧光参数变化
2.5 镉胁迫对桢楠幼树吸收光能分配的影响
桢楠幼树叶片对吸收光能的分配情况如表4所示。正常生长条件下,桢楠幼树叶片吸收的能量约70%均用于热耗散,约20%用于非光化学反应,约4%用于光化学反应,说明桢楠幼树叶片对吸收光能的利用率较低,多通过热耗散途径耗散,以免光抑制。而Cd胁迫进一步减弱了桢楠幼树叶片对吸收光能的利用能力,且在T2及以上处理时,天线热耗散能量减小,过剩激发能增加,表明PSⅡ光化学反应中心受损,保护调节作用减弱。

表5 不同水平镉处理下桢楠幼树叶片对吸收光能的耗散情况
3 讨论和结论
3.1 镉胁迫对桢楠幼树叶绿素含量的影响
叶绿体色素是植物进行光合作用的基础物质,其含量变化能反映植物光合作用的潜力。其中Chla、Chlb作为天线色素,在光能的吸收和传递中占主导作用,而Car除具备天线色素功能外,还可直接猝灭单线态氧,减小活性氧数量,以减轻逆境下植物所受的膜脂过氧化损伤。本研究中三者含量均呈下降趋势,这可能是Cd与叶绿体中多种酶的巯基(—SH)结合或取代叶绿体的主要组成成分如Fe2+、Zn2+和Mg等,破坏叶绿体结构,加快了叶绿素的分解[7,25]。此外,我们在研究镉对桢楠幼树生长及抗性生理的影响(已另文发表)时发现细胞分裂素(CTK)含量有所减少,这也可能是导致叶绿素含量减少的一个重要原因[26]。当植物叶绿素含量不足时,若想维持较高的光合速率,便会调节Chla/b值,本研究中Chla/b值逐渐升高,应是桢楠对镉毒害的一种防御机制[27],此外有研究表明,减少捕光叶绿素蛋白(Chlb)的合成数量,可以降低活性氧的积累[28],增强自身的耐性。
3.2 镉胁迫对桢楠幼树气体交换参数的影响
Pn、Tr、Ci、Gs能够反映植物光合生理对逆境的响应情况,是评价光合作用强度的基本指标。其中Pn变化的原因主要包括气孔因素和非气孔因素两种[29],且根据Faqruhar和sharkeyt[30]的观点,判断叶片光合速率降低的主要原因是气孔因素还是非气孔因素,要依据两个数值的变化方向,即Ci和LS;且Ci变化是主要判断依据。本研究中,随着胁迫程度的加重,Pn逐渐降低,Ci增加,LS降低,表明Cd胁迫环境中,限制桢楠幼树光合速率的主要原因是非气孔因素,这也与董袁媛等[27]研究镉胁迫对黄麻光合作用的影响,谭长强等[31]研究镉胁迫对蚬木光合生理的影响,所得到的结果相似。而引起Pn降低的原因可能较多,如叶片叶绿素含量不足、遭受光抑制,导致PSI和PSII的电子传递速降低等,这与简敏菲等的研究结果一致[25],也与本文在叶绿素含量、PSII方面的研究结论相符。
3.3 镉胁迫对桢楠幼树叶片光合-光响应特征的影响
桢楠幼树的Pn均随着光合有效辐射的增强而逐渐增大,直至稳定,这与干旱、水淹胁迫下桢楠幼树光合-光响应特征一致[32- 33]。许大全提出,AQY和Pmax同时下降是植物遭受光抑制的显著特征[34]。本研究中,AQY、Pmax值逐渐降低,可见Cd胁迫下桢楠幼树除对弱光的利用能力降低以外[16,35],强光及饱和光强下其光合速率也会受到明显抑制。另外,Fv/Fm值变化也能衡量植物遭受光抑制程度[17,34],本研究中,其值在50 mg/kg时显著降低(表4),结合AQY、Pmax和Fv/Fm值,可以认为,在Cd胁迫环境下桢楠幼树遭受到一定程度的光抑制。而LSP降低,LCP升高则体现出桢楠幼树对光抑制的适应能力,这与滕振宁等人的研究结果一致[36]。
3.4 镉胁迫对桢楠幼树叶绿素荧光参数的影响
叶绿素荧光作为PSⅡ光化学反应的指示器,对分析植物光合作用功能有重要作用。研究表明,Cd胁迫能抑制PSⅡ光合活性以及电子运转[37]。Fv/Fm、Fv/F0在T1—T4处理时变化不明显,这说明T1—T4处理时PSII反应中心受损程度较小,桢楠幼树仍能维持一定的光能转换效率以及光合活性,随着Cd浓度的进一步增加(T5),Fv/Fm、Fv/F0显著下降,表明PSⅡ综合功能会有所下降,吴坤等[26]认为这与叶绿体超微结构发生改变有关。ΦPSⅡ和ETR均能反映植物在PSⅡ的实际光化学反应效率,本研究中,二者值均显著下降,这与徐小逊等[[2]和苏秀荣等[39]的研究结果相似,Cd胁迫导致桢楠幼树叶片的光合电子传递链受到破坏,电子无法有效从PSⅡ向PSⅠ传递,同时PSⅡ反应中心也无法接受新的光子,一旦光子累积过多,叶片的细胞同化力(NADPH和ATP)则无法正常形成,C的固定和同化也无法完成,ΦPSⅡ和ETR也就因此下降,桢楠幼树光合速率降低的原因可能也在于此,这与Vassilev和Yordanov[40]的研究结果一致。qP(光化学淬灭系数)能反映PSⅡ中天线色素吸收的光能参与光化学反应的比例。本研究中qP显著下降,吴坤等认为这是植物处于Cd胁迫环境时,为防止自身PSⅡ原始电子受体QA被过度还原的适应现象[26],同时植物也会利用叶黄素循环途径将过量光能猝灭耗散(NPQ上升),以降低自身光合机构的受损度[41]。
3.5 镉胁迫对桢楠幼树吸收光能分配的影响
植物叶片所吸收的光能通常包括3部分:天线热耗散的能量(D)、光化学耗散能量(P)和非光化学反应耗散能量(Ex)[22,42]。本研究中,桢楠幼树以天线热耗散途径为主来消耗过剩光能,Cd胁迫下光化学耗散能量减小,这与光合、荧光参数(AQY、Pmax、Fv/Fm)降低有关;而非光化学反应耗散能量的增加可防止PSII和电子传递链被过分还原,进而避免过剩光能对光合机构的过度破坏,这反映了桢楠幼树对Cd胁迫的适应能力。
综上所述,低浓度(T1—T2)Cd胁迫时,桢楠幼树叶绿素含量、气体交换参数和光响应特征参数就明显下降,随着Cd浓度的进一步增加(T3—T5),桢楠幼树PSⅡ反应中心遭受一定程度的光抑制,对弱光以及吸收光的利用能力变弱,最终导致桢楠幼树光合能力下降,因此不宜在镉污染较为严重的地区大量发展桢楠人工幼林。
