云南文山石漠化区车桑子叶脉密度与叶氮含量关系对生境的响应
2020-07-17庄立会沐建华周建松
陈 静,庄立会,沐建华,周建松
文山学院环境与资源学院,文山 663099
叶脉与叶氮含量是描述叶经济谱中相互联系、协同变化的两个重要性状,共同揭示了植物功能属性间的内在关联[1],反映植物在自然压力下选择的生态对策,是理解植物环境可塑性机制的关键途径[2]。叶脉网络散布于叶片系统之中,形成叶脉密度、大小、间距和闭合度等功能性结构,叶脉密度是叶脉系统中最关键的指标,体现叶脉与表皮气孔、叶肉细胞的的接触程度,在很大程度上决定了叶片的光合产物、水分和养分等物质运输效率与安全性[3],与叶片光合性状的关系密切。叶氮含量作为植物的固有属性,是解释叶经济谱的重要生化参数[4],常用测量指标包括单位质量叶氮含量(Nmass)和单位面积的叶氮含量(Narea),其中Nmass高低直接决定植物叶片光合能力的大小[5],Narea则为Nmass与比叶面积之比,反映了单位叶面积的潜在光合能力[6]。叶脉主要由木质部细胞和韧皮部细胞组成,氮素的输送数量和氮素在叶片结构中分配比例取决于叶木质部和韧皮部的组织数量及其运输能力[7],由此表明,叶脉密度与Nmass和Narea联系紧密,可反映植物叶片网络系统的构建策略[8]。
在土壤贫瘠的石漠化地区,土壤环境物理特征及辐射状况的改变,诱导着各种生态因子发生变化,并形成一定的环境梯度[9],进而影响到植株生理功能结构的优化。光照、土层厚度异质性营造出差异化的局部小生境,使种群的资源获取和利用受到限制,植株面临着选择压力的风险,为了自身的生态适合度,倾向于调节叶脉密度和叶氮含量的投资比例,尽量延长叶片氮素的存留时间,达到提高叶氮利用效率与光合能力的目的[10],实现不同生境中植物对叶片氮分配的控制。所以,研究石漠化区植物叶脉密度与叶氮含量关系的生境响应,有助于深入了解植物维持营养平衡的叶脉可塑性机制。
车桑子(Dodonaeaviscosa)又名坡柳,为热带和亚热带常绿灌木,无患子科车桑子属植物,耐旱力强、萌生率高,是喀斯特石漠化区的先锋树种,在水源涵养、水土保持、维持区域生态平衡等众多方面具有极其突出的生态功能。近年来,国内外学者围绕植物叶脉密度与叶片功能性状的关系进行了大量的研究,较为系统的阐述了不同环境下叶脉密度的构建机制。韩玲等[11]对张掖湿地芨芨草的研究显示,植物叶大小与叶脉密度呈显著的负相关,宋丽清等[12]比较了高粱和紫苏的叶脉密度与光合特性,提出叶脉密度与光合速率呈显著的正相关;Huang等[13]通过对光照和黑暗条件下中国台湾南部C4植物的研究发现,高叶脉密度的发育需要提高生长素的生物合成和运输,Baresch等对巴拿马和美国马里兰州不同光环境下森林植物叶片结构的探究表明,解剖和发育上的限制有助于被子植物获得最高的叶脉密度、气孔密度和生理速率[14]。目前对车桑子的研究多集中在植被恢复过程中的种群特性[15]、气候因子对生长发育的影响[16]、海拔对叶片功能性状[17]和土壤化学计量特征的影响[18]及不同居群的抗旱机理[19]等方面,但车桑子叶片化学计量特征和叶脉性状的生态学意义的研究不够深入,尤其石漠化区不同生境车桑子叶脉密度与Nmass和Narea关联性的研究还鲜有报道。本文通过探索云南文山石漠化区不同群落生境下车桑子叶脉密度和叶氮含量的变化,旨在理解先锋植物叶片生物物理构建与化学性状的关系,揭示石漠化地区叶片碳水代谢与营养代谢间的投资机理。
1 研究地区和研究方法
1.1 研究区概况
研究区位于云南文山东山乡石漠化区(23°54′N,104°58′E),海拔1315—1788 m。年均温16.11℃,年均降水量989.30 mm左右,年均蒸发量1780.20 mm,相对无霜期309d。土壤以赤红壤和红色石灰土为主,pH值4.5—5.5。植被分布兼具水平和垂直分异特征,以陆生乔灌林木为主,主要有车桑子(Dodonaeaviscosa)、茶条木(Delavayatoxocarpa)、清香木(Pistaciaweinmannifolia)、白刺花(Sophoradavidii)、小叶鼠李(Rhamnusserpyllifolia)、青刺果(Prinsepiautilis)、火棘(Pyracanthafortuneana)、云南松(Pinusyunnanensis)、墨西哥柏(Cupressuslusitanica)、旱冬瓜(Alnusnepalensis)等植物。
1.2 样地设置与调查方法
于2018年8月10日—20日完成样方调查。基于群落和环境调查,在研究区选择海拔和坡度基本相同,坡向不同的旷地(I)、林缘(II)、林下(III)3个自然样地,经主成分分析发现,第一主成分主要反映土壤物理状况和光照条件,其中土层厚度是最关键的主导因子,而光合有效辐射(Photosynthetically active radiation,PAR)是最重要的限制因子,第二主成分主要反映了土壤全氮含量,所以PAR、土层厚度、土壤全氮是生境样地最主要的3个影响因子,进而导致空气温湿度和土壤含水量的差异(表1、表2)。旷地生境样地位于南面,光合有效辐射和气温较高,但郁闭度、空气湿度和土壤水分均较低,林下生境位于北面,郁闭度、空气湿度和土壤水分均较高,而光合有效辐射和气温偏低,林缘样地地处西偏南,其生境状况界于旷地和林下之间。在3种生境样地分别设置10个10 m×10 m样方,总计30个样方,然后对每个样方选取3株车桑子(大中小各1株),共90株,在其植冠中部外侧东、南、西、北4个方位各标记成熟向阳3片叶片,共12片,然后进行以下步骤。
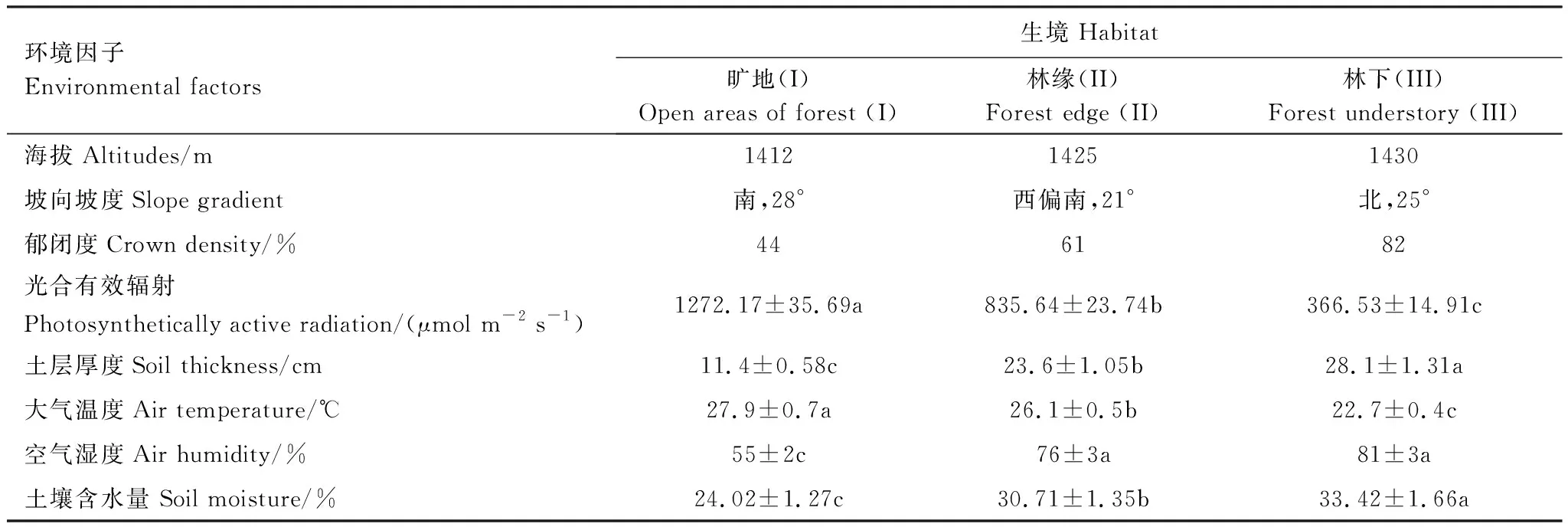
表1 不同生境车桑子群落的环境因子
1.3 实验方法
1.3.1环境特征和光合参数测定
选择3个晴朗天气的6: 00—18: 00,用便携式光合仪(LCpro+,ADC BioScientific Ltd.,UK)进行3个生境的光合有效辐射、大气温度、空气湿度等气候因子的测定,并同步测定并记录净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)等光合参数,每2 h观测1次,全天共测定6次,结果取平均值;然后在各个样地用钢钎法进行土层厚度的测量,同时采集土壤样品,重复3次,带回实验室测定土壤含水量;土壤全氮含量(mg/g)采用全自动凯氏定氮仪(KJELTEC TM 8400,FOSS Ltd.,Denmark)测定。光能利用率(Light use efficiency,LUE)(%) =Pn/PAR、水分利用效率(Water use efficiency,WUE)(μmol/mmol) =Pn/Tr。
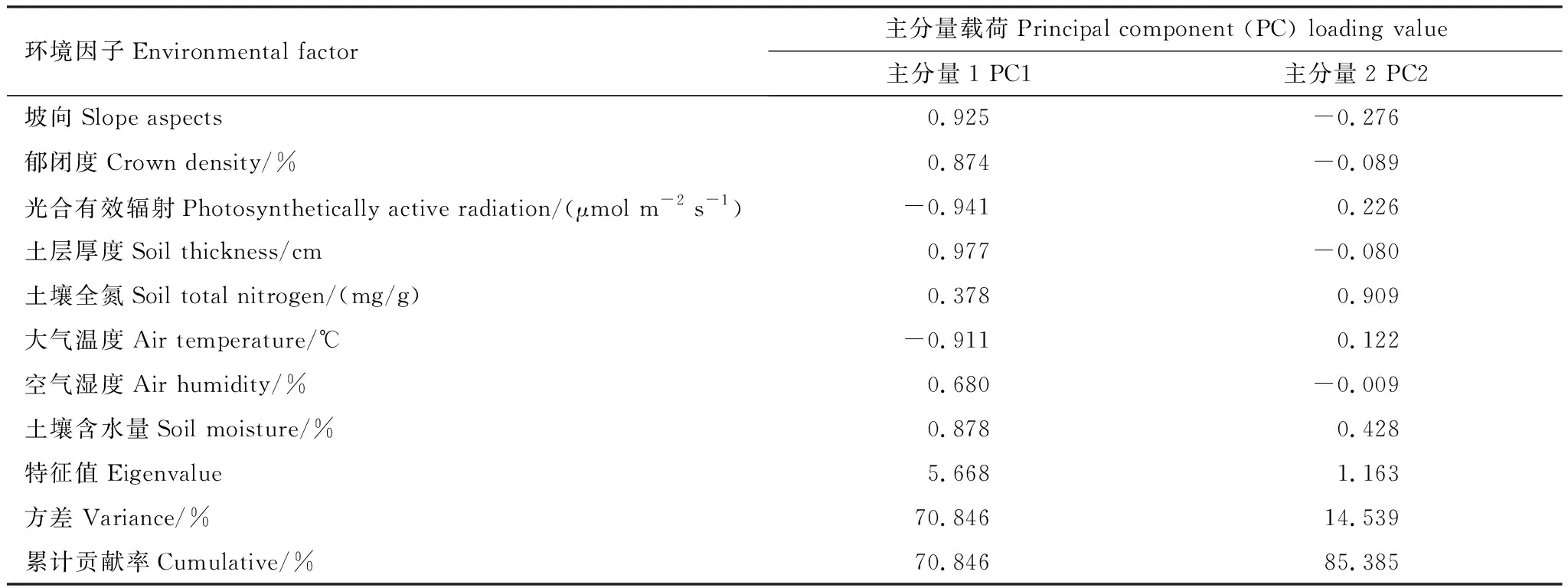
表2 不同环境因子的主成分分析
光响应曲线测定时间为9: 00—11: 00,通过LCpro+的光控制系统,设定14个PAR梯度(0、50、100、150、200、400、600、800、1000、1200、1400、1600、1800、2000 μmol m-2s-1),测定其瞬时光合速率,3次重复,计算平均值。以PAR为横轴,光合速率为纵轴绘制光合作用光响应曲线拟合光合作用光响应曲线方程,推算光饱和点(Light saturation point,LSP)和光补偿点(Light compensation point,LCP)。
1.3.2叶脉密度测定
选取标记过的所有叶片,用福尔马林-乙酸溶液固定后,通过5%NaOH的乙醇进行化学清理,并用番红-固绿染色,水包埋使之呈透明薄膜状在体视显微镜(SMZ168-BL,Motic,中国香港)下观测叶脉密度。选择3个视野拍摄每个叶片的两端及中间部位,采取Motic Images Plus 2.0软件获得各个照片中叶脉的总长度,总叶脉密度(mm/mm2) = 叶脉总长度/单位叶面积。
1.3.3比叶面积和叶绿素含量测定
用便携式激光叶面积仪(CI- 202,CID,Camas,USA)测定进行每个标记叶片的叶面积,最后将叶片烘干至恒质量,测定叶干重,比叶面积SLA(cm2/g) = 叶面积/叶干重。叶绿素含量采用丙酮提取法,使用分光光度计(UV1700,南京科捷,中国)测定。
1.3.4叶氮含量测定
将所采叶片烘干至恒重后研碎,利用Vario EL III型元素分析仪(Elementar Analysensysteme GmbH,Hanau,Germany)测定单位质量叶氮含量Nmass(mg/g)[20],并计算单位面积叶氮含量Narea(g/m2) =Nmass× 10/SLA[21],光合氮利用效率(Photosynthetic nitrogen use efficiency,PNUE)(μmol mol-1s-1) =Pn/Narea[22]。
1.4 数据处理方法
本研究中所有原始数据全部采用Microsoft Excel 2013整理。3个生境梯度旱柳植株性状平均值的差异比较采用单因素方差分析(α= 0.05);然后利用线性回归的方法对不同生境下车桑子叶脉密度和叶氮含量的关系进行研究。数据统计和绘图分别由SPSS 20.0及oringin 8.5软件完成。
2 结果与分析
2.1 不同生境车桑子光合参数的变化
如表3所示,比叶面积、叶绿素总含量、光饱和点、光补偿点、蒸腾速率、气孔导度、水分利用效率和光能利用效率在3个生境的差异显著(P<0.05);净光合速率在林缘(II)与旷地(I)和林下(III)之间具有显著差异(P<0.05),而在I和III间表现不明显(P> 0.05)。从旷地依次到林缘、林下生境,比叶面积、叶绿素总含量、光能利用效率呈递增趋势,其中林缘(II)较旷地(I)分别上升16.51%、14.95%和92.45%,林下(III)较林缘(II)分别上升21.12%、15.61%和58.82%;光饱和点、光补偿点、水分利用效率呈递减趋势,林缘(II)车桑子较旷地(I)分别减小了11.68%、25.90%和25.57%,林下(III)较林缘(II)分别减小了36.52%、27.93%和13.76%;车桑子净光合速率、蒸腾速率、气孔导度呈先升后降的倒U型趋势,在林缘生境(II)均具有最大值。表明叶片的光合性状有较高的敏感性和可塑性,能够适应于不同生境。

表3 不同生境车桑子光合参数(平均值±标准差)
2.2 不同生境车桑子叶脉密度与叶氮含量的变化
3个生境车桑子的叶脉密度与叶氮含量存在显著差异(P< 0.05,表4)。随着旷地(I)转到林缘(II)、林下(III),车桑子叶脉密度(Vein density,VD)、Narea均呈明显的递减趋势,林缘(II)比旷地(I)分别减少了16.20%、6.11%,林下(III)比林缘(II)分别减少了8.25%、8.84%;Nmass呈明显的递增趋势,林缘(II)比旷地(I)增大了9.59%,林下(III)比林缘(II)增大了10.45%。可见,VD与Narea存在一致性的变化规律,而与Nmass有着相反的变化趋势。

表4 不同生境车桑子叶脉密度与叶氮含量(平均值±标准差)
2.3 不同生境车桑子叶脉密度与单位质量叶氮含量的关系
3个生境车桑子VD和Nmass的关系如图1所示。车桑子VD和Nmass在旷地(I)和林下样地(III)均呈显著负相关关系(P<0.05),在林缘样地(II)二者的负相关关系不显著(P>0.05),从生境I依次到III,方程斜率绝对值逐渐减小。表明随光照强度递减,车桑子VD减小,Nmass增加,与II和III相比,生境I植物对VD的资源投入比例大于Nmass;生境I和III车桑子VD和Nmass之间形成相互制约的权衡关系,生境II是车桑子VD和Nmass变化的过渡区域,VD和Nmass呈不显著的权衡关系。
2.4 不同生境车桑子叶脉密度与单位面积叶氮含量的关系
不同生境车桑子VD和Narea的关系如图2所示。车桑子VD和Narea在旷地(I)和林下样地(III)均呈显著正相关关系(P<0.05),在林缘样地(II)二者的正相关关系不显著(P>0.05),从生境I依次到III,方程斜率逐渐增加。表明VD越大,Narea也越大,在某一给定的VD时,车桑子样地I比II和III对Narea的资源投入更大;生境I和III的车桑子VD和Narea之间具有显著的协同关系,生境II是车桑子VD和Narea由大到小的过渡区域,VD和Narea之间呈不显著的协调性。
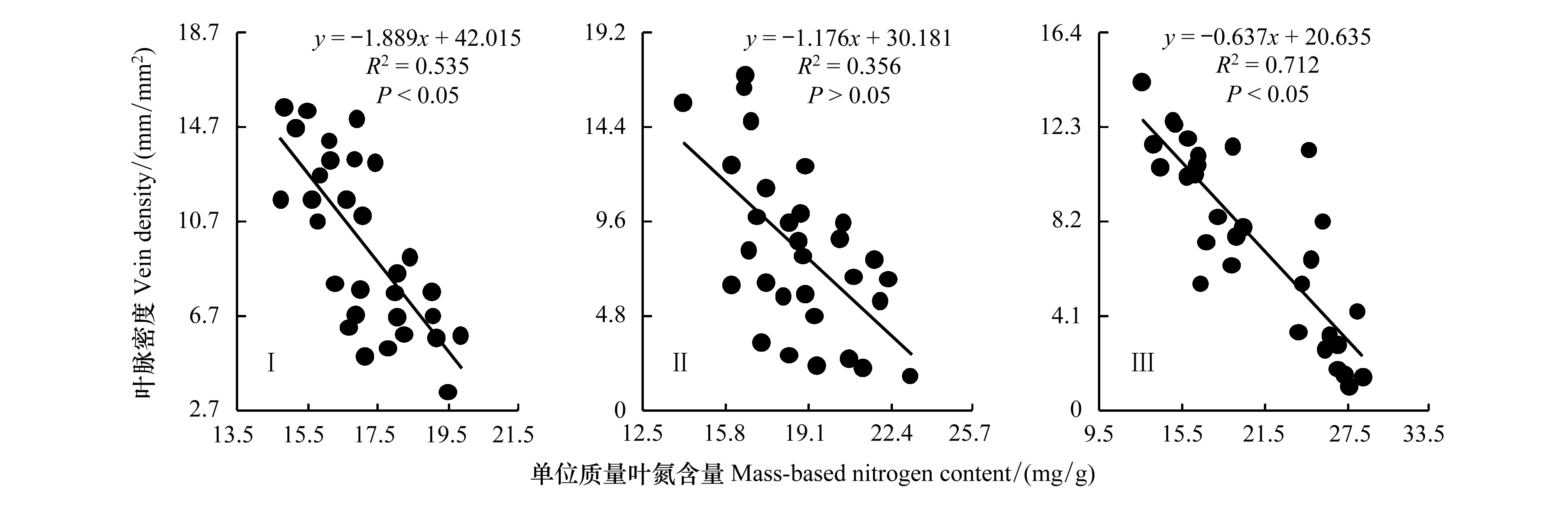
图1 不同生境车桑子叶脉密度与单位质量叶氮含量的的关系Fig.1 Relationship between vein density and Nmass of Dodonaea viscose in different habitats

图2 不同生境车桑子叶脉密度与单位面积叶氮含量的关系Fig.2 Relationship between vein density and Narea of Dodonaea viscosa in different habitats
3 讨论
长期处于异质环境下的同种植物形态发育和构型功能会产生不同的响应,产生趋异适应[23]。车桑子在不同生境条件下构建最佳的叶脉网络性状,从而保证叶片正常的氮素分配与平衡。研究区属于亚热带喀斯特石漠化生态系统,具有低纬太阳辐射强烈,土层浅薄,岩石裸露等特征。本研究表明,PAR、土层厚度与土壤全氮是不同生境条件下车桑子群落的直接影响因素,随着旷地生境转向林下生境,车桑子VD、Narea呈明显的递减趋势,Nmass呈明显的递增趋势;在石漠化区旷地和林下生境,车桑子VD与Nmass和Narea分别具有显著的负相关(P<0.05)和正相关(P<0.05),在林缘生境,VD与Nmass和Narea分别呈不显著负相关(P>0.05)和正相关(P>0.05)。支持了Ordoez等[24]关于“植物叶结构建成的资源投入越多,维持光合作用功能的投资就越少”的观点。但与朱济友等[25]的“叶脉密度与单位质量叶氮含量呈极显著正相关”观点完全相反,这源于前人是以人工绿化林为研究对象,本文基于石漠化山地自然林的研究,在没有人为干扰的状态下,通过植物叶片尺度上的经济谱规律,能够更准确地解释叶性状与生态环境的耦合机制。
VD受外界环境因子的影响明显[26],其变异性反映了植物对环境的可塑性响应及异质性资源利用策略。在3个生境梯度,PAR和温度对植物叶脉性状起着重要作用,VD的变化为:旷地 > 林缘 > 林下。旷地生境PAR平均值接近于叶片光饱和点,午间光照强烈,常超出光饱和点,PSII的最大光化学转化效率下降会引起光抑制现象,同时高温严重影响了光合碳同化过程,造成了叶内酶活性失灵,间接导致植物的光抑制[27],强光和高温成为植物生长的主要限制因子,车桑子的光合器官形成多种保护机制来最大程度地减少光热可能带来的潜在伤害,一方面车桑子形成较厚的角质层和紧密的栅栏组织[18]减少光透射率来降低LUE,选择厚叶片以抵御强光对植物的损伤;另一方面增加叶脉维管束组织的构建投入,确保叶木质部和韧皮部管道运输路径的安全性,维持从叶基部向叶肉细胞的水分传输功能,减轻高温胁迫对植物的伤害效应[28],故叶片倾向于具有高VD。由旷地过渡到林缘,群落光照和热量资源从冗余到适宜,车桑子拥有相对优势的种群生态位,环境因子对植物生长的限制较小,无需改变叶脉网络结构也可获得最高的Pn,车桑子选择中等VD。林下的种间和种内邻体干扰效应明显增强,遮阴现象严重,植物面临着弱光胁迫,PAR大幅度下降引起的温度降低会导致车桑子生长激素(如ABA等)水平升高,调控基因机制,进而阻止细小叶脉的发育[29],车桑子增加叶绿体的体积和叶绿素含量,扩大细胞接受太阳辐射的光合面积,提高LUE以补偿低VD造成的光能损失。
Nmass对土壤营养状况的依赖度较高,土壤有机质及氮含量直接影响植物体内氮素含量的高低[30],其多寡决定叶的建成消耗[31],从而影响植物叶脉的构建。在旷地生境条件下,低的土壤全氮会导致植物体内的氮素供应不足,降低了参加光合作用的酶含量与酶活性[32],车桑子将相当一部分氮素给予保卫组织、结构物质和贮藏物质的构建以抵御氮胁迫,抑制了到达叶片内部叶绿体的光强度、加大了CO2在叶肉组织中的传导阻力和拉大了气体在叶内的传导距离,降低了叶氮向光合器官的分配比例[33],这种将有限的氮素资源更多用于叶片防御组织的构建模式,使得Nmass下降而VD上升。林缘生境具备良好的土壤水氮条件,保证了植物的水分运输效率[34],为了满足植物生长所需的氮素供给,车桑子充分发挥光合量子驱动作用,加强叶片内部氮营养循环与调节,叶片光合系统氮分配比例与细胞壁氮分配比例较为均衡,最终促成适中的Nmass和VD。林下生境的土壤全氮含量较高,利于其增加叶片捕光组织的氮分配比例,提高光截获能力,与此同时较低的光补偿点有助于提高叶绿素含量和碳氮物质的积累,更多的氮被用于光合作用结构的构建[35],从而导致分配到单位质量的叶氮比例增加,减少了叶脉的构建成本,表现出高Nmass低VD。
Narea综合表征了Nmass和SLA的变化,由于SLA变幅大于Nmass,所以SLA可决定Narea大小,高SLA代表低Narea[36]。Narea和VD与WUE均具有显著关联性[36- 37],因而Narea与VD存在一定的联系。土层厚度是影响土壤储水能力的重要方面,植物的生长状况在很大程度上取决于土层厚度[38]。然而旷地生境的土层厚度较薄,土壤含水量较低,为减少植物水分亏缺,车桑子通过增加细胞壁厚度加大水蒸气进出表皮阻力[39],显著降低了Gs和Tr,提高了WUE。叶片厚度随细胞壁厚度变化而变化,致使叶片变厚[40],SLA减小而Narea增大;同时厚的细胞壁意味着木质部气蚀化程度较高,栓塞风险增大,车桑子叶片需要增加维管数量以绕过气蚀化的木质部把水分运输到蒸腾位点,从而形成发达的叶片维管组织,VD由此增加,高VD增强了植物对干旱贫瘠环境的耐受能力,但也降低了植物的生长速率[41]。从旷地转向林缘、林下生境,相对较厚的土层增加了土壤储水能力和土壤水分对植被的承载能力[42],促进了SLA的增加,而使得Narea下降。拥有中等SLA的林缘车桑子叶片,适度降低叶肉组织与叶脉的接触面积,选择适中的VD最大限度协调自身水分运输与外界光能获取之间的关系,以获取最大的光合收益;具有高SLA的薄叶,以牺牲WUE和VD为代价增大水分传输速率,减小用于构建防护结构的氮碳投资[43],从而使林下车桑子有着较高的相对生长速率。
VD、Nmass和Narea作为叶经济谱的衡量指标,三者间的协同性与权衡性反映了先锋物种在石漠化生境的存活机理。位于旷地的车桑子具有最大的VD和Narea,最小的Nmass,形成少氮多脉的低SLA叶片,因而VD与Nmass和Narea分别存在显著的负相关和正相关,表明叶片营养物质在保卫结构方面的投资较多,偏于“保守型”厚叶的一端,属于高消耗慢收益的“缓慢投资-收益”型[44],这归因于旷地植物需要长时期积累物质以满足自身的生长发育;林下车桑子具有最小的VD和Narea,最大的Nmass,具有多氮少脉的高SLA叶片,VD与Nmass和Narea的相关性相同于旷地生境,表明叶片在光合结构方面的投资相对较多,靠近“竞争型”薄叶的一端,倾向于低投入快生长的“快速投资-收益”型[44],原因是林下植物需要加快生长速率以适应林分层片间的竞争[45]。林缘车桑子Pn和Tr均达到最高,较强的光合能力使其可以同时兼顾植株WUE和LUE等功能需求,车桑子不必遵循叶经济谱的资源权衡策略,采取折中VD和叶氮含量的均衡适应模式,最大限度扩大生态优势,维护种群的稳定。因此,林缘车桑子的VD和叶氮含量的相关性未达到显著水平。
4 结论
车桑子叶脉密度与叶氮含量的关系,揭示了石漠化区植物叶结构和叶片化学性状之间的资源分配权衡机理。本研究发现,随着光照条件和土壤状况的改变,车桑子群落生境产生了明显差异,植株通过调整叶经济谱“投资-收益”型来响应异质性生境。生长在旷地的车桑子倾向于构建高叶脉密度,采取较小单位质量叶氮含量和较大单位面积叶氮含量的“缓慢投资-收益型”的保守策略,形成低比叶面积的厚叶片以适应强光和干旱环境;而生长在林下的车桑子利用良好的土壤条件,选择低叶脉密度、高单位质量叶氮含量和低单位面积叶氮含量的“快速投资-收益型”的竞争策略,实现资源的最大化利用以适应弱光胁迫的环境。自然选择压力使不同生境的车桑子找到最适的投资与收益关系,以最“经济”的方式构建叶脉网络与营养功能结构,较好地验证了石漠化区植物适应异质生境的快速反应机制。
