科尔沁沙质草地生物量积累过程对降水变化的响应模拟
2020-07-17王明明刘新平何玉惠车力木格孙姗姗
王明明,刘新平,何玉惠,魏 静,车力木格,孙姗姗
1 中国科学院西北生态环境资源研究院奈曼沙漠化研究站, 兰州 730000 2 中国科学院大学,北京 100049 3 中国科学院西北生态环境资源研究院乌拉特荒漠草原研究站, 兰州 730000 4 中国科学院西北生态环境资源研究院皋兰生态与农业综合研究站, 兰州 730000 5 北京大学深圳研究生院人居环境科学与技术重点实验室, 深圳 518000
全球气候模式预测未来降水格局将会发生显著变化,极端降水和极端干旱发生频率增加,极端干旱和湿润年份比例上升,降水量、降水发生频率、降水季节分配模式以及大小雨比例等都会有不同幅度的变化[1- 4]。降水格局的变化对陆地生态系统有很强的影响,从次降水事件特征的变化到年际间的降水的波动都会改变生态系统水分可利用性以及养分周转,从而对生态系统的结构和功能产生影响[4-5]。
科尔沁沙地作为中国北方最严重的沙漠化区域之一[6-7],在沙漠化过程中沙地原生植被退化严重,生态环境脆弱,极易受到气候变化和人类活动的影响[8]。科尔沁沙地气候为半干旱陆地季风气候,降水主要以小降水(<5 mm)事件为主,且降水变异性强[9],近50年来的气象资料统计分析发现科尔沁地区气候有暖干化趋势,这为该区域的沙漠化防治带来了更大的挑战[10-11]。水分是干旱生态系统最主要的限制性因子,其中许多关键的生态过程受水分变化的影响强烈[12- 15],尤其是植被生物量积累与降水变化密切相关,因此理解植被生物量积累过程对降水变化的响应对科尔沁沙地沙漠化防治及生态恢复具有重要意义。目前在科尔沁沙地植被生物量与降水变化之间关系的研究主要集中在较大空间尺度上降水特征变化对生态系统生产力的影响[16- 18],尽管在一些典型区域也开展了一些野外观测实验,主要研究植被与土壤水分以及地下水之间的关系[19],但对年内植被生物量积累过程对降水变化的响应机制仍不清楚。另外,关于科尔沁沙地植被与降水变化关系的研究大多数只关注降水量和植被特征之间的关系,但在实际的生态系统中降水首先要转化为土壤水分才能被植物所利用,因此将植被、降水以及土壤水分结合起来是研究植被对降水变化响应的关键。另外,这方面的研究目前大多关注较大空间尺度上植被生物量的年、季节或月的变化及其与降水之间的关系[20- 24],而对点尺度下土壤水分和植被生长的相互作用研究匮乏。本研究中基于植被-土壤水相互作用的耦合方程,结合植被对降水事件的延迟-阈值响应模式,构建模型来研究植被生物量积累对降水变化的响应。利用本模型可以在点尺度上实现植被生物量积累过程的日尺度模拟,这有助于在更加精细的时空尺度上理解植被与降水之间的关系,这对沙漠化土地生态恢复以及后续经营管理具有重要的意义。
1 研究方法
1.1 建立模型
1.1.1植被生长和土壤水分动态耦合模型
在半干旱沙质草地中水分是影响植被生长最主要的环境因子,土壤水分变化在一定程度上决定着植被的存活、生长及演替,而植被的生长也对土壤水分动态以及水分可利用有重要影响,二者相互作用,耦合变化。在本研究区降水是土壤水分最主要的补给源,而蒸散和渗漏是两个主要的土壤水分消耗过程,因此,土壤水分的动态变化主要由这3个过程的综合作用决定。对植被生长而言,光合作用是生物量积累途径,植物自身呼吸、茎叶脱落以及根系死亡分解是生物量消耗的过程,这些过程的综合作用导致植被生物量的生长季动态变化。基于上述变化过程,生长季土壤水分变化和植被生物量积累过程可用1式表示[25]:
(1)
式中,B和s分别表示t时刻的植被生物量和土壤水分相对含量,A为植物光合同化速率,R为植物自身呼吸速率,Q为植物叶片脱落及细根分解速率,I为降水量,E为蒸散量,D为渗漏量。
在受水分的限制的生态系统中土壤水分是影响植被光合速率以蒸散发的主要的因子,因此可以假设当s>sw(sw为植被凋萎时的土壤相对含水率)时叶片气孔完全开放,当s (2) 式中,Am是日潜在同化量。同样地,日蒸散量可以由潜在蒸散量(Em)和土壤水分(s)表示: (3) 式中,Em为日潜在蒸散量,ka和ke是当ED为Em一半时的s-sw的值,因为AD和ED的都随着气孔的开闭而变化,所以认为ka=ke,统一为“半饱和”系数k。 将2式和3式带入1式,再将生物量和土壤水分的变化统一到单位面积的样方上,引入植被功能性状参数和土壤水力参数后可以得到如下方程: (4) 式中,ρ为比叶面积,fL为叶干物质含量,Yg为同化效率,Rr为根呼吸系数,q为机械损耗速率(茎叶脱落和细根死亡等),n是土壤孔隙度,Zr为植被根系深度,Ks为土壤饱和导水率,c为关于土壤类型的常数,KsSc代表深层渗漏。将方程中nZr除到右边,并设γ=ρfLEm/nZr,α=AmρfLYg,β=Rr(1-fL)+q,变换后得到5式,其中γ的实际意义为单位生物量的最大蒸腾速率,α为最大同化速率,即为相对生长速率,β为由根系呼吸、细根枝叶死亡脱落造成的损失速率。 (5) 1.1.2关键参数计算 化简后模型中存在三个关键参数α、β和γ,这三个参数实际上反应了潜在的蒸散发速率、光合同化速率以及植被生物量损耗速率。有研究表明在水分匮缺的生态系统中植物光合速率的变化对降水脉冲极为敏感,一次有效降水事件的发生会引起植被光合速率的脉冲式响应[26- 28]。对此Reynold等在重新评估“水分分割理论”和“脉冲贮存理论”后,综合这两种理论建立了植被对降水脉冲的阈值-迟滞响应模型(Threshold-Delay, T-D),主要解决单次有效降水事件如何转化为植被响应的问题。该模型中设定植被对降水事件的响应存在最高(RU)和最低(RL)的降水阈值,在此范围内的降水为有效降水。当前的响应变量可由上一时刻的响应变量和当前的增量表示[29],即: αt=km×αt-1+δt (6) 式中,αt为t时刻的植被某一过程的相对变化速率,km为降水脉冲过后αt的衰减系数,δt为αt在t时刻的响应增长,计算如下: (7) (8) 式中,αmax为α的最大值,δ*为δ的最大潜在增加量,δmax为最大响应增长,τ为响应时滞。植被光合同化速率受降水脉冲的响应显著,因此用T-D模型进行计算,而β和γ受降水脉冲的影响不是很大,因此在模型中设定为常数进行计算。 生物量对降水变化的响应计算如下: (9) 式中,Bchange为降水变化下模拟的最大生物量,Bunchange为实际降水条件下模拟的最大生物量。 1.1.3模型参数厘定 科尔沁沙地位于半干旱区,其沙质草地生态系统在很大程度上受水分的限制,因此适合于该模型假设条件。模型实测参数均在位于科尔沁沙地的中国科学院奈曼沙漠化研究站周边的典型的沙质草地测定。模型参数如表1所示,其中导水指数q是一个经验常数,根据土壤类型设定[30]。根区土壤孔隙度n和饱和导水率Ks使用环刀分层(0—10 cm,10—20 cm以及20—40 cm)取样,在实验室每个深度进行3次重复测定,最终取平均值。最大日蒸散量(Em)和根系呼吸速率(Rr)参考已有文献[31],蒸散和同化速率半饱和系数(k)参考文献[32]。群落平均比叶面积(ρ)和叶干物质含量(fL)参考文献[33-34]。T-D模型参数RU和RL通过查阅文献获取[35-36],ymax、δmax、τ参考Reynold等测定结果,km由实测数据拟合得到。 1.2.1研究区概况 研究区位于内蒙古自治区科尔沁沙地(42.92°N, 120.70°E),属于半干旱陆地季风气候,夏季高温,冬季寒冷。年均温为6.4 ℃,大于10 ℃的年积温在3000 ℃以上,无霜期约150 d。年降水量360 mm,75%的降水发生在6—9月,年蒸发量在1600—400 mm之间,4—9月蒸发量占全年蒸发量的78%左右。土壤类型主要有风沙土、草甸土、沙质栗钙土和沼泽土[37]。地貌以固定沙丘、半固定沙丘、流动沙丘和面积不等的平缓沙地和低洼地交错分布为特征。根据奈曼沙漠化研究站2005—2018年间未发表的观测数据,该区域地下水平均埋深12—15 m。植被斑块化分布和空间异质性较强,植被盖度较小的流动沙丘上一年生先锋物种沙蓬为优势种,半固定和固定沙丘上差不嘎蒿(Artemisiahalodendron)和小叶锦鸡儿(Caraganamicro-phylla)等灌木为优势种。盖度较高的沙质草地以菊科一二年生物种黄蒿(Artemisiascoparia)以及多年的芦苇(Phragmitesaustralis)和白草(Pennisetumflaccidum)为优势物种[38]。 表1 模型参数及其取值 1.2.2数据获取及处理 气象和降水数据采集于中国科学院奈曼沙漠化研究站自动气象站。沙质草地样方生物量数据采集于2013年、2015年和2017年的4—9月中旬,其中地上生物量采用收割法,随机选择6个50 cm×50 cm的样方,剪下活体生物量,带回实验室烘干测定生物量干重;地下生物量用直径10 cm的根钻按0—10 cm、10—20 cm和20—30 cm的深度分层取样,每个样方3个重复,同一深度组成混合样,装入0.5 mm的网袋中用流水冲洗后剔除死根,将活根在80 ℃的恒温下烘至48 h后称量,计算样方内地下生物量。总生物量为地上、地下部分之和,单位取g/m2。模型求解和绘图均在MATLAB 2016完成。 科尔沁沙质草地一般从每年的4月中下旬开始返青,生物量积累开始,9月份生长停止,植被枯黄,生物量开始降低。沙质草地3年的生物量实际监测结果如图1所示,生物量的增长整体上呈现单峰模式,即在生长季第150(设定4月1日为第1天)天左右达到最大,之后逐渐下降。模型模拟的生物量积累过程与实测的植被生物量积累趋势基本一致,也呈现出单峰模式,满足本研究分析降水变化下植被生物量积累响应的需求。生长季初期植被生物量积累缓慢,随着降水的逐渐增多,生物量积累出现明显的跃增。土壤水分模拟结果显示土壤水分变化对降水事件的发生存在明显的响应,随着降水事件的发生,土壤水分出现明显的脉冲式增长,降水过后开始下降,但这种响应随着植被生物量的积累而逐渐减弱,反应出了植被对土壤水分动态的调控作用。 图1 生物量积累过程以及土壤水分模拟结果Fig.1 Simulation of biomass accumulation and soil moisture实线为模拟生物量积累结果,黑圆点为实测结果生物量 降水量变化直接影响半干旱草地初级生产力,在2016年的降水基础上通过改变单次降水量,保持降水发生频率、时间以及季节分配不变,模拟降水量变化对植被生物量积累过程的影响。结果如图2所示,降水量增加,促进植被生物量累积,降水量减少,抑制植被生物量累积。根系层土壤水受降水增加的影响不大,这主要与沙地高的饱和导水率有关;降水量减少,土壤水分也相应地降低减少。为了进一步量化生物量积累对降水量变化的响应大小,在2013至2017年5生长季的降水基础上进行改变,保持其他降水特征不变,分别改变增减降水量0%、10%、20%、40%,每个变化幅度下分别随机计算1000次,计算生物量的响应,发现生物量对降水量增加和减少的响应存在显著差异,生物量对降水增加的响应远大于对降水量减少的响应,整体上表现出明显的非对称响应,这与野外原位控制实验的观测结果相一致[39, 40]。 图2 不同降雨量下的植被生物量积累过程、土壤水分变化及响应 (-表示降水量减少,+表示降水量增加)Fig.2 Response of vegetable biomass accumulation for the change of precipitation amount (-, The decrease of precipitation; +, The increase of precipitation) 降水频率的变化直接改变土壤湿润模式,进而影响土壤水分的可利用性。不同降水量下降水频率的改变可能会对植被生物量积累过程产生不同的影响,这里选择1个典型的湿润年份(2016年)和1个干旱年份(2009年),在这两年的基础上保持降水总量不变(图3),通过随机减少降水次数来模拟不同降水量基础下的降水频率变化对植被生物量积累过程的影响。经过1000个生长季的随机模拟后计算不同降水频率下植被生物量的响应,结果如图3所示。可以发现湿润年和干旱年生物量积累对降水频率变化的响应存在很大的差异,干旱年降水频率的增加对植被生物量积累有明显的负效应,随着降水频率增加幅度越大,这种负效应逐渐加强;而在湿润年降水频率增加对植被生物量的积累存在正效应,当降水频率增加幅度为40%时正效应达到最强。生物量积累对降水频率减少的响应在干旱年和湿润年基本相同,一定变化幅度内,降水频率降低抑制生物量的积累,但是当超过某一阈值后促进生物量积累,这个阈值在干旱年位于40%—60%的减少幅度之间,而在湿润年位于20%—40%的之间;当降水频率降低幅度达到60%时生物量对频率变化的响应突然变大。 图3 湿润年和干旱年生物量积累对降水频率变化的响应Fig.3 Response of vegetation biomass accumulation for changes on precipitation frequency in wet and dry year 植被-土壤水分耦合模型是基于植被生物量积累速率方程和土壤水分平衡方程来描述植被与土壤水分之间的耦合关系,模型中降水作为植被生物量和土壤水分变化的驱动因子,其变化直接引起植被生物量积累和土壤水量平衡的变化。该模型虽然形式简单,但可以直观地反应植被和土壤水分的关系,并且通过改变降水特征,可以模拟的生物量和土壤水分对降水变化的响应。Zea-Cabrera等应用该模型研究了不同植物间水分竞争的“公地悲剧”现象,Nordbotten等在该模型的基础上引入降水的随机特征,推导出了植被生物量的随机概率分布,阐述稳条件下的植被和土壤水分之间的动态关系[25, 41],但在上述研究中都设定植被相对生长速率α恒定。然而,许多研究已经发现植被相对生长速率受许多因素的影响,尤其在受水分限制的生态系统中其与降水脉冲密切相关[42- 44]。T-D模型基于植被对降水脉冲的阈值迟滞响应模式,引入植被功能类型参数和生理参数,实现了对受水分限制的植被在脉冲降水下相对生长速率的模拟,从理论上完善了植被对降水脉冲的响应模式。引入T-D模型来计算降水波动下的模型参数α,实现了土壤-植被耦合模型中相对生长速率的时变模拟,使得该模型更加接近植被实际的生长过程。 模拟结果表明降水量增加显著促进植被生物量积累,降水减少抑制植被生物量积累,但是植被对降水量变化的响应存在明显的非对称性。我们的研究中降水是土壤水分唯一的补给源,通过改变单次降水量来模拟降水量的变化,这种变化下单次降水量的减少会使更高比例的降水成为无效降水,而单次降水的增加会使更高比例的降水成为有效降水,有效降水量不成比例的变化使植被生物量积累对降水量变化也表现出非线性响应。在实际观测中也发现植被生物量对降水变化存在明显的非对称响应[45],这表明我们的模拟结果能较好的反映植被生物量积累对降水量变化的响应。植被生物量对降水量变化的非对称性响应与植物在不同水分条件下物质分配规律以及群落稳定性有关。一般而言,湿润条件下植物会分配更多的光合产物到地上部分去捕获更多的光,从而提高生产力,而在干旱环境中分配更多的光合产物到地下部分去获取更多的水分和养分,提高自身的竞争力。不同的水分条件下植被地上和地下比例的变化在一定程度上会导致了生物量积累对降水量变化的非线性响应[39]。另外,植物群落对环境变化具有一定的抵抗力,如植物群落中不同功能型的物种在降水量发生变化时会产生互补效应,进而维持群落生物量稳定性[46-47]。植物特殊的功能性状和生理特性也是引起这种非线性响应的主要的原因,降水量发生变化时这些植物会通过调节自身的植被功能性状以及生理过程来适应降水的变化,如科尔沁沙地一年生草本猪毛菜(Salsolacollina)会根据降水变化调整物候来适应降水的变化,完成生命周期[39]。根系层土壤水分在降水量减少时显著降低,而在降水增加时变化不大,这主要是因为当降水量增加后根系层土壤水分大多时候处于盈余状态,使得更多的土壤水分向深层转移,根系层土壤水分含量实际上变化不大,相反降水量减少时土壤水分亏缺,受降水量变化的影响较大。 保持降水总量不变,改变降水频率会使单次降水量、降水间隔以及有效降水量都发生变化,进而使得土壤水分可利用性发生改变,导致植被生物量积累过程产生不同的响应[48]。将1000次随机改变降水频率后的植被生物量响应平均后发现,有效累积降水量变化率与生物量响应在干旱年和湿润年相关性都很微弱;而单次有效降水(>5 mm的降水事件)与生物量响应在干旱年和湿润年都呈显著正相关因此单次有效降水量的大小是影响植被生物量对降水频率变化响应的关键因素;有效降水间隔(相邻两次有效降水的时间差)在干旱年与生物量响应呈显著正相关,在湿润年与生物量响应无关;累计有效降水量在干旱年和湿润年都与生物量响应不存在显著关系(图4)。根据等级响应理论[49],在响应阈值范围内植物对降水脉冲事件的响应存在等级效应,T-D模型设定植物对降水脉冲的响应也遵循等级响应理论,在响应范围内植物的相对生长速率对降水事件的大小有很强的依赖性,因此单次有效降水量的大小是影响植被对降水变化响应首要因素。虽然累计有效降水是许多单次有效降水事件的加和,但降水频率变化引起的降水间隔的变化会抵消生物量地累计有效降水的影响,因此其整体上对生物量的影响不大。Heisler[44]等发现也发现增加单次降水的大小,有利于地上生物量的积累,这与本研究的模拟结果基本一致。 图4 有效累积降水量变化率和累计有效降水间隔与植被生物量响应之间的关系Fig.4 Relationship between effective cumulative precipitation change rate and cumulative effective precipitation interval with vegetation biomass response上排为干旱年,下排为湿润年,a、d单次有效降水量变化,b、e有效降水间隔变化率,c、f为有效累积降水总量变化率 模拟结果还显示生物量对降水频率变化的响应存在一定的阈值,且响应阈值在湿润年和干旱年并不一致,这主要与干旱年和湿润年降水特征有关。干旱年降水量和降水次数都少于湿润年,降水频率增加使单次有效降水量减少,而在湿润年有效降水次数较多,单次有效降水量较大,降水频率增加使得单次有效降水量降低的同时缩短了有效降水间隔,有效降水间隔的缩短抵消了单次有效降水减少的效应,二者之间的这种相互抵消在增加幅度为40%的时候达到最强,因此在湿润年降水频率增加促进生物量积累,而在干旱年抑制生物量积累。类似地,降水频率减少会增加单次有效降水,但同时也延长了有效降水间隔,因此存在对降水频率变化的响应阈值,且该阈值在因降水量和降水特征的不同而不同。相比于2009年,2016有许多大于20 mm的降水,因此随着降水频率的增加,会使超过20 mm的部分无效降水转化为有效降水,但同时也会使部分降水变成小于5 mm的无效降水,当这两种变化的正效应最强时达到响应的阈值;2009年超过20 mm的降水较少,降水频率增加引起的大降水转化为有效降水的部分不足于抵消有些降水变成小于5 mm降水的损失,因此降水频率增加抑制生物量积累。总的而言,降水频率的变化通过改变植被单次有效降水量和有效降水间隔来影响生物量的响应。 植被生物量积累过程对降水变化存在很强的响应,且在不同的降水变化下存在不同的响应模式。(1) 植被-土壤水分耦合模型和T-D模型结合能准确模拟半干旱沙质草地点尺度植被生物量累积过程,模型模拟结果与实测生物量累积趋势基本一致。(2) 植被生物量对降水量变化表现出明显的非线性响应。模拟结果显示降水量增加,促进植被生物量积累,反之,抑制植被生物量积累,但生物量积累对降水量增加的响应程度远大于降水量减少,整体上,生物量积累对降水变化的响应表现出很强的非线性。(3) 生物量对降水频率变化在不同的降水特征下表现出不同的响应模式,干旱年降水频率增加抑制生物量积累,湿润年则促进生物量积累,且增加40%左右时促进效应最强;降水频率在一定减少幅度内抑制生物量积累,当超过一定阈值后反而促进,但该阈值在干旱年位于40%—60%的减少幅度之间,而在湿润年位于20%—40%的减少幅度之间。降水频率变化引起的单次有效降水量变化是引起生物量积累响应的关键因素,有效降水间隔的变化在干旱年也对生物量积累的响应有很大的影响。1.2 研究区概况及数据处理

2 结果分析
2.1 植被生物量积累过模拟

2.2 降水量变化对植被积累过程的影响

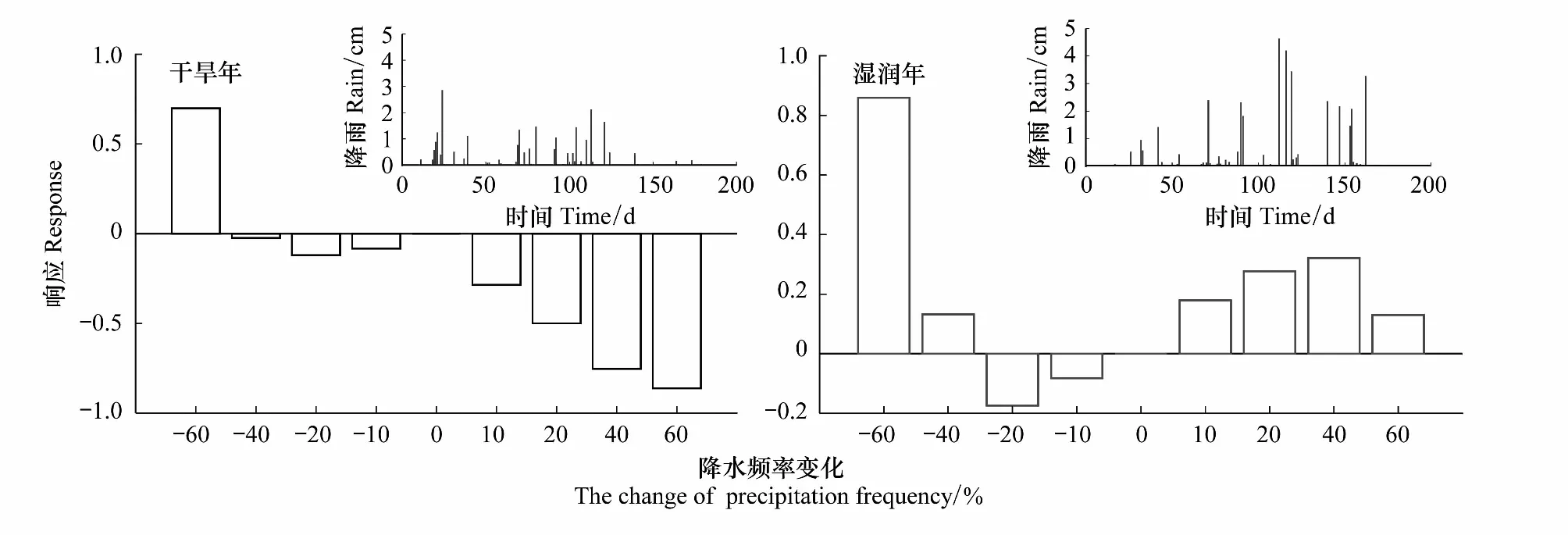
2.3 植被生物量积累对降水频率变化的响应
3 讨论
3.1 模型有效性
3.2 生物量积累对降水量变化的非线性响应
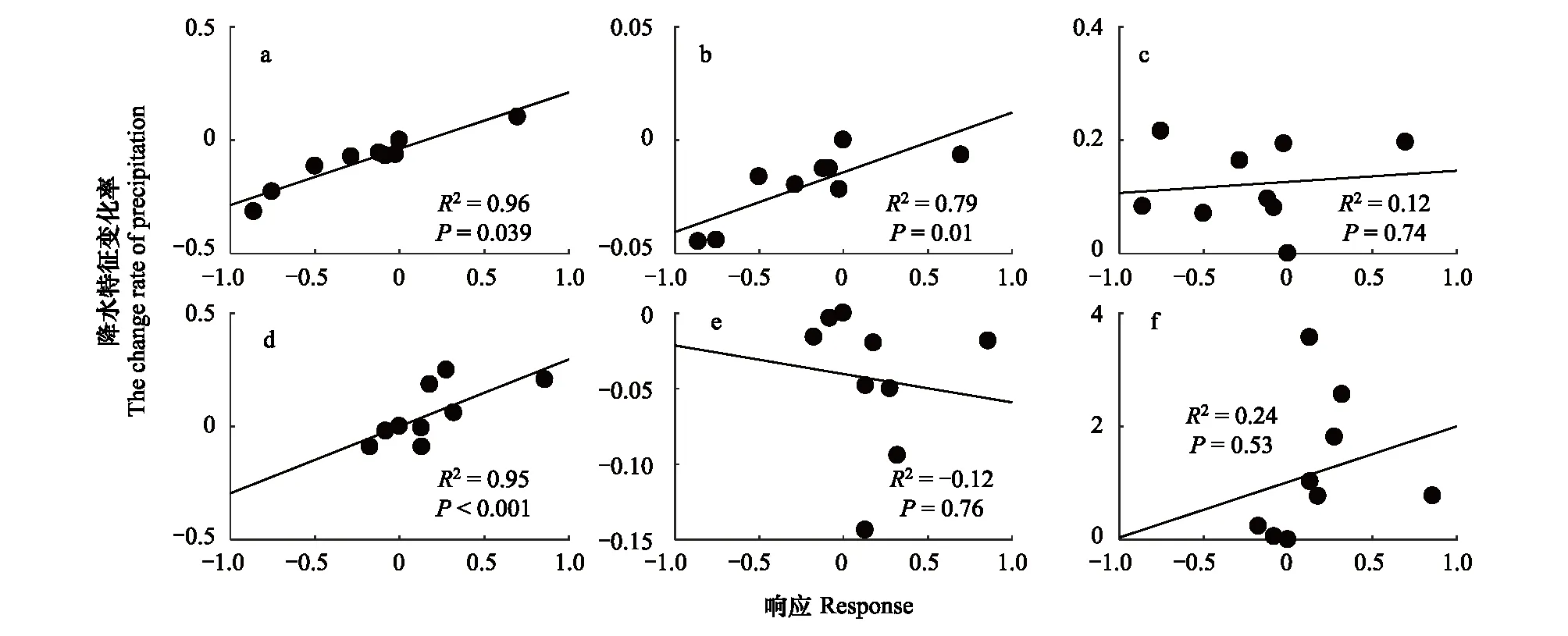
3.3 生物量积累对降水频率变化的响应
4 结论
