植物叶寿命测定方法综述
2020-07-16任海燕韩国栋
朱 毅,任海燕,韩国栋
(内蒙古农业大学草原与资源环境学院/草地资源教育部重点实验室/农业部饲草栽培、加工与高效利用重点实验室/内蒙古自治区草地管理与利用重点实验室,内蒙古呼和浩特 010018)
叶片在自然界以各式各样的形态存在,其最大的作用在于通过光合作用固定碳,这也是植物获取碳的最主要途径。Wright等人基于全球植物性状网络数据库,研究了叶片性状之间的关系,确立了叶片经济学谱(Leaf economic spectrum),即表征叶片生产性状(如光合速率、氮磷含量)与表征叶片维持和消耗性状(如叶寿命、比叶重)之间存在权衡关系,并认为这是植物在长期进化中与环境适应的结果[1,2]。例如,光合能力强的叶片往往由于对防御方面的投资较少而寿命较短[3]。全球变化导致资源供给改变,进而影响植物对资源(水分、养分等)的获取能力,然而在资源获取过程中植物面临着效率和寿命的权衡[4]。因此,叶寿命直接影响植物对资源的获取和利用,是反映植物物候的重要指标,连接着叶片生理生态、植株个体生长和生态系统过程,它是影响生态系统养分循环和生产力的决定性因子[4~6]。叶寿命是指叶片个体或叶片部分存活和生理活动的时间段,它以天、月或年表示[7]。有研究表明,从叶片出生开始,随着叶面积的增加到完全展开状态,光合速率也随之增加至最大,随后随着叶片的衰老逐渐下降[8,9]。叶寿命的研究具有极其重要的理论价值和应用前景。
1 叶寿命研究历程
植物叶寿命的研究最早可追溯到1914年,Harper首次报道了叶寿命与生境养分可利用性相关[10]。随后到了20世纪60~80年代,更多的研究集中在叶寿命的生态适应性方面,尤其是在落叶和常绿生境中。较有代表性的如Monk于1966年在Ecology上发表的关于常绿生境的生态学意义[11]。1982年,Chabot和Hicks更详细地阐述了叶寿命的生态学意义[8]。Coley等人1985年发表在Science上的文章,研究了资源可利用性、植物抗草食性防御和叶寿命的关系,发现叶寿命取决于资源可利用性,影响植物不同周转率防御的相对优势[12]。
上世纪90年代,对叶寿命的研究达到了高潮,更多的学者投入到叶寿命的研究中来,在各种生境下开展叶寿命的研究工作,主要探讨叶寿命与植物碳收益、养分动态、植物防御和地理分布的关系。代表性的学者如日本的Kikuzawa[13],基于模型从植物的碳获得和碳消耗之间的权衡关系出发,研究了叶寿命变异的原因及意义;美国的Peter Reich等[14]则从叶片水平、植株个体水平和立地水平对叶寿命展开了研究;西班牙的Escudero等[15]发现,植物具有长寿命的叶片是植物提高养分利用效率的适应性表现;瑞士学者Diemer[16]首次通过灌木的叶寿命数据集阐述了叶寿命的生物地理分布格局;Craine等[17]则首次利用叶寿命模型测定了14种植物(6种禾本科、8种杂类草)的叶寿命。到21世纪,植物功能性状再次成为全球热点研究问题,而叶寿命作为最重要的功能性状之一也备受关注。在全球变化的背景下,叶寿命与其他功能性状的关系以及尺度推演方面的研究尤为突出,代表性学者有Wright、Shipley和Reich等人。Wright等人首次在全球尺度上阐明了6个关键叶片性状之间的相互关系,重点分析了LL-LMA(叶寿命-比叶重)这组关系[1];之后,Bill Shipley等人基于全球数据集,通过模型讨论了LL、Am(最大光合速率)、Nm(单位质量叶片氮含量)、LMA之间的权衡关系[18];Ren等从植物激素乙烯的角度,揭示了荒漠草原植物叶寿命随氮素添加变化的生理机制[19];Xu等在Kikuzawa(1991)研究的基础上,改进了性状驱动的碳最优模型,预测了潮湿热带林的叶寿命变化,揭示了叶寿命的变异是如何由叶片结构特征和非生物环境约束下的碳优化产生的[20]。
尽管叶寿命的相关研究已有一个多世纪(表1),但目前仍然存在许多问题:如缺少公认的测定叶寿命的方法;不同叶片性状之间的权衡关系制约着对叶寿命变异机制的客观评价;全球变化对叶寿命影响的研究较少等。

表1 植物叶寿命研究热点文献
2 叶寿命的测定方法
叶寿命是指植物叶片从出生到衰老的时间长度。一般把完全展叶定义为叶片的出生,部分或完全枯黄定义为叶片的死亡[17]。叶寿命的测定方法主要包括生命表法、传统的野外标记观测方法和野外监测与叶寿命模型结合的方法。其中,编制生命表的方法一般用来测定常绿阔叶树种的叶寿命,与人口统计学中的生命表类似,计算的是叶片的期望寿命,而与人口统计中的生命表不同的是,叶寿命的生命表包含的主要指标包括叶片的龄级、存活叶片数、脱落叶片数、总存留叶片数以及叶片的期望寿命。标记观测法,就是选取一定数量的植物个体,对其叶片进行标记并进行观测,观测间隔时间一般较短,类似于物候观测,最后经过大量重复样本的综合分析得出某种植物的叶寿命。标记观测法包括传统的野外标记观测法以及后来基于标记观测发展的标记观测与叶寿命模型结合的方法。与传统的标记观测方法相比,采用标记观测与叶寿命模型结合的方法具有减少工作量的优点,计算的结果为某物种叶片的平均寿命,能够更好的比较禾本科和其他不同生长形态的植物的叶寿命。
2.1 生命表法
编制生命表的方法一般用来测算常绿阔叶植物的叶寿命。由于常绿阔叶植物的生活期较复杂,叶寿命往往大于1,有的甚至可以存活3~4个生长季。为了能够精确的推算常绿阔叶植物的叶寿命,可以根据枝条上的芽鳞痕所形成的节环以及枝条的粗细程度、颜色等特征来识别枝条的生长年龄,并对不同年龄枝条上的叶片数计数。以1年为一个年龄级,规定当年形成的叶为1龄叶,去年形成的为2龄叶,以此类推,然后据此编制成生命表(见表2)。计算叶片的期望寿命,公式如下[21,22]:
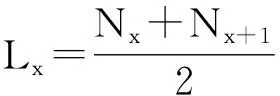
式中:x为叶龄级(按年划分等级);Nx为在x年龄级开始时的存活叶片数;Dx=Nx-Nx+1,即x到x+1年龄期的脱落叶片数;Lx为从x到x+1年龄期的平均存活叶片数;Tx为龄期x及其以后各年龄级的叶片留存总年数;Ex为x期开始时的平均期望寿命。
注:引自宋永昌,2000。
由于表中各项数据具有统计关系,只要测得同龄出生的叶片总数Nx,各龄级开始的存活叶片数Nx或该龄级期限内的脱落叶片数Dx,即可通过上述公式计算出其他数值。Tx与Lx是专为计算Ex而设立的,所以在生命表的各个参数中,只有Nx或Dx是直接观测值,其余的都是统计值。
采用编制生命表测定叶寿命的方法主要适用于对常绿阔叶树种的研究。例如:王希华等通过生命表法对天童国家森林公园的常绿阔叶树种叶寿命进行了研究,发现常绿阔叶树种的叶寿命大多在1~3年之间,小乔木及灌木的叶寿命要大于中乔木[22];田玉鹏等使用生命表法对福建梅花山51种常绿阔叶植物进行了研究,发现灌木的叶寿命最长,而小乔木的叶寿命最短,并且同种植物的叶寿命随海拔升高而延长[23],这可能是环境因素的差异造成两个研究结果的不同,是植物适应环境的一种表现;王瑞雪等采用生命表法对江苏宝华山常绿植物的虫食频度进行研究发现,叶寿命越长虫食频度也越高[24]。
2.2 传统的野外标记观测方法
采用野外标记观测法测定叶寿命的方法虽然劳动强度大,但以重复监测时间间隔跟踪植物个体叶片出生和死亡无疑是最好的方法。具体做法是随机选取植物个体,抽取部分叶片并挂上小标签进行标记,定期记录它们的存活或死亡情况,间隔时间大约为预估叶寿命的1/10。而为了提高精确性,每个物种至少需要40个叶片,最好是能达到160个叶片。为了达到这个数目,建议增加物种的个体抽样数,而不是增加每个植物个体的枝条数量。根据长期监测得到的数据,计算每个叶片的寿命,最后取每株的平均值[7]。如Sydes曾报道了对石灰岩草地进行叶群体统计[25],对于要测定叶寿命的物种,随机选取一株植物个体作为代表,标记抽样的叶片,并每间隔一段时间进行监测,记录叶片存活的叶长,当叶片上没有任何活的绿色组织时便认为叶片死亡,叶片的出生率和死亡率为自上次抽样日期以前每个植物每天出现或死亡的叶数。在确认得到的结果近似于服从正态分布以后,采用t-检验对观测样本进行验证分析:
这种测定叶寿命的方法缺点是工作量太大,需要对植物叶片以重复间隔长期观测,并且对于叶寿命的估计是整个叶片而不是单位叶长(单位叶长更能代表生物量周转率),因此在比较禾本科和其他不同生长形态的物种叶寿命时可能不够准确。
2.3 标记观测与叶寿命模型结合的方法
因为以上野外标记观测法存在的局限性,Craine & Reich等人提出了一种新的将野外标记观测与叶寿命模型结合的测定方法[17,26]。此方法和上述标记观测法类似,当一片叶子被认为完全展开时,此时被定为它的“出生时间”,当叶片至少有2/3变成黄色或棕色时,则被认为死亡。选取一部分植物完全展开叶片进行标记,长期监测被标记叶片的叶面积或长度。该方法认为,在第一次监测时已经有了一些植物叶片,虽然出生时间不同,但都会在监测期间死亡。监测开始后,出生较早的一部分叶片会陆续死亡,而一些出生较晚的叶片在检测结束后还未死亡。减去这两部分后就是在监测期间出生并死亡累积的叶面积,再减去累积死亡的叶面积就是在寿命期内的累积叶面积,除以叶片面积后就得到叶寿命。用此方法测出来的是某物种叶片的平均寿命。
根据模型,平均叶寿命(LL,day)等于在监测期间产生并死亡的累积叶片长度(m·day)或面积(m2·day)除以叶片长度(m)或面积(m2),计算公式如下:
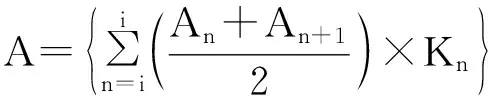
式中:i表示第一次监测时间;j表示最后一次监测时间;An表示从开始监测到n天之间累积产生的叶片长度或面积;Sn表示从开始监测到n天之间累积死亡的叶片长度或面积;Kn表示监测时间n和n+1之间的间隔;ns表示当累积死亡的叶片长度或面积等于第一次监测时的叶片长度或面积的累积量时(即S=Ai)的时间;nl表示当累积产生的叶片长度或面积等于最后一次监测死亡的叶片长度或面积的累积量时(即A=Sj)的时间。
如图1[17]:曲线A(方形)表示在整个监测期间产生或死亡的叶片累积长度或面积(上述式A);曲线B(三角形)表示整个监测期间死亡的叶片累积长度或面积(上述式B);曲线下面区域的面积是产生或死亡叶片长度或面积的总天数(A下面区域)和产生叶片长度或面积的总天数(B下面区域),这两条曲线之差表示未死亡的叶片长度或面积的天数。区域C表示在监测开始时就已经存在(出生日期未知)的叶片长度或面积(即方程式C);区域D表示在监测结束还没有死亡(死亡日期未知)的叶片长度或面积(即方程式D)。从曲线A和B之间的区域减去C和D(A-B-C-D),就得到监测期间产生并死亡的叶片累积长度或面积(m·day或m2·day,即区域E)。将区域E除以区域E中的总叶片长度或面积(m或m2,区域E的高),就得到叶片的寿命,相当于区域E的水平长度。该模型的机理类似积温,如果在知道某个时期的积温(℃·day)和这一时期的平均温度(℃)时,将积温除以平均温度就可以得到该时期的天数(day)。
在采用这种新的将标记观测和模型结合的方法测定了14种植物的叶寿命之后,Craine & Reich随后又使用了同样的方法对10种草原植物展开研究,发现叶寿命在提高CO2的供给下延长了3.4d,而在提高N供给的条件下缩短了4.2d[26]。Ren等(2011)使用该方法对我国内蒙古典型草原的5个重要物种进行原位研究发现,不论在丰雨年还是贫雨年,水分添加减少了叶寿命约10d,氮素添加减小了叶寿命约5d[27]。此外,Ren等(2013)采用此方法测定盆栽和大田实验中的植物叶寿命,通过设置不同梯度的氮素添加实验(5、10、15 g N m-2yr-1),表明氮素添加增加植物乙烯生产,降低叶寿命;同时乙烯生物合成抑制剂(CoCl2)的添加减小乙烯生产,增加叶寿命;揭示氮素添加通过影响植物激素乙烯而导致叶寿命的变化[19]。该研究首次从植物生理角度为已经广泛存在的生态学现象(即氮输入影响叶寿命)提供了全新的解释。
为什么植物叶寿命对养分以及CO2的供给变化做出如此响应,许多专家学者对于叶寿命的变异做了大量的研究,其中“碳平衡”理论被广泛接受,它认为叶寿命是植物优化碳收益的结果[8]。植物为了最大限度的提高净碳收益,当叶片每单位时间的净碳收益在其生命周期内达到最大值时,叶片必须更换,这就是最佳叶寿命理论。最佳叶寿命理论认为,植物的叶片以及支撑叶的相关茎、根等其他一切组织的建造成本和这些器官及组织的后期维护成本,共同决定了植物的最佳叶寿命,从而能使植物获得最大净碳收益。基于这种假设,日本学者Kikuzawa提出了最佳叶寿命模型[13]:
topt=[2bc/(a-m)]1/2
式中:topt表示最佳叶寿命;b为光合速率降低为0所需要的时间;C表示叶片建成消耗;a表示某一时刻的瞬时光合速率;m为a时刻的瞬时呼吸速率。从式中可以看出,影响叶寿命的因素为叶片净光合速率(a-m)、叶片建成消耗(c)以及叶片光合作用降低速率(1/b),由此可以得到以下推论:叶寿命随叶片建成消耗的增加而变长;叶寿命随光合作用降低速率的增加而变短;叶寿命随最大光合速率的增加而变短。
虽然此模型在一定程度上能够通过叶片净光合速率来预测植物的叶寿命,但由于在建模时植物的功能性状相互作用常常被忽略,叶片建造成本被低估等因素,限制了模型的预测能力。对此,Xu等人优化了该模型[20]:
式中:G为平均碳收益率;t为叶龄;CCleaf为叶片层级的叶片建成生物化学消耗;CCplant为没有被计入的植物层级消耗;LMA为比叶重,视为常数,等于成熟叶片的LMA。该模型的核心思想是优化叶片整个生命期内的平均净碳收益。最佳叶寿命被定义为G(t)达到最大值时的时间,也就是G(t)和g(t)相等时的时间。
Xu等人的研究结果表明,植物可以根据光的可利用性动态调整叶片生物化学特性以达到最佳碳收益[20],Lloyed等人的研究也支持了这一结果[28]。Kikuzawa等从碳收益的角度研究了叶寿命的全球模式,将f定义为一年中每月气温和水分可利用性有利的部分,在短f的条件下,常绿植物需要较长的叶寿命才能获得最大的碳收益,而落叶植物的叶寿命随着f的缩短而降低[29]。有报道表明,生长在贫瘠土壤和生长在阴凉处的植物叶寿命更长,可能是不利的环境因素限制了植物的光合作用,而植物则通过延长叶寿命来获得更大的碳收益[30~33]。当然,植物不会无限制的延长叶寿命,根据Xu等人的研究,叶寿命回报(Leaf lifetime return)虽然随着叶寿命的增加而增加,但碳收益的价值可能会随着时间的延长而降低[20,34]。叶寿命长的好处只有在叶片实际能够达到最佳叶寿命时才能实现,各种环境不利因素都能在叶片达到最佳寿命之前杀死它们,而短叶寿命的叶片潜在损伤率可以降低。
综上所述,叶寿命的测定方法主要有生命表法、标记观测法、标记观测与叶寿命模型结合法三种,其中基于观测芽鳞痕的编制生命表的方法只适用于针对常绿阔叶树的研究,此方法已被广泛使用,国内已有许多应用此方法的报道[22~24];而传统的标记观测的方法适用于针对乔木以及草本植物的研究,有着十分可靠的优点,但很明显的缺点是工作量太大;采用标记观测法和叶寿命模型结合的方法同样适用于乔木及草本的研究,这种方法虽然在可靠性上比不上传统的标记观测法,但在保证可靠性的同时可大大的缩减工作量,尤其在对草原植物的研究时是最为推荐使用的一种方法。几种不同方法的优缺点比较详见表3。

表3 叶寿命测定方法
3 结语
从上世纪初一直到本世纪,针对叶寿命的研究一直在开展。因为叶寿命具有非常重要的生态学意义,它连接着叶片生理生态到植株个体生长和生态系统过程。在叶片水平,叶寿命与叶片氮含量、光合作用和氮素转移息息相关;在植株水平,叶寿命对植物净碳固定和氮素吸收起重要作用;在生态系统水平,叶寿命是生态系统净初级生产力和氮素循环的一个重要决定因子。加强叶寿命及其相关性状的研究,可以为当前的生物地球化学循环模型提供重要的科学依据。虽然国外学者已经有大量关于叶寿命研究的报道,但多数集中在对于常绿或落叶树种及其在全球尺度上的地带性研究[20,29],而对于草地植物及其对资源环境供给的变化的响应方研究甚少,特别是在中国非常缺乏这方面的数据,因此我们现在很有必要开展相关方面的研究。虽然对于叶寿命的研究已经长达近一个世纪,但对于叶寿命的测定一直缺少一种公认的研究方法。此外,叶寿命及其相关性状在陆地生态系统模型的研究中还未引起足够的重视,需要更多的研究来弥补这些缺口。
