北京油松上东京伞滑刃线虫的鉴定
2020-07-15刘乐乐方亦午陈先锋顾建锋
刘乐乐, 方亦午, 刘 曦, 陈先锋, 边 勇, 顾建锋
(1.宁波中盛产品检测有限公司,浙江 宁波 315100;2.宁波海关技术中心/宁波检验检疫科学技术研究院,浙江 宁波 315100;3.北京市林业保护站,北京 100029;4.北京海关技术中心,北京 101300)
伞滑刃线虫属(Bursaphelenchus)是1937年由Fuchs[1]提出并建立,目前已报道的有效种多达120种,该属线虫主要习居于树木中,大多靠取食真菌为生.其中,松材线虫[B.xylophilus(Steiner & Buhrer,1934) Nickle, 1970]和椰子红环腐线虫[B.cocophilus(Cobb, 1919) Baujard, 1989]具有较强的致病性,且是国际重要检疫对象[2].
在我国,松材线虫于1982年在南京中山陵的黑松上首次被发现[3],之后逐步扩散.据国家林业和草原局2019年第4号公告,我国松材线虫疫区已达18个省588个县级行政区,病死松树超过1 000万株.目前,该虫向北已扩散到辽宁省铁岭市,红松(PinuskoraiensisSieb. et Zucc.)和落叶松(LarixolgensisHenry)为新确认的松材线虫自然感病寄主[4-5].
2019年初,北京市林业保护站在对北京市周边林区的松材线虫进行普查时,发现一株油松(PinustabulaeformisCarrière, 1867)有枯萎病症,取病木样送至北京海关技术中心进行分离,获得大量的伞滑刃属线虫,最后宁波海关技术中心将其鉴定为东京伞滑刃线虫(BursaphelenchustokyoensisKanzaki, 2009).
据报道,东京伞滑刃线虫仅在日本东京的赤松(PinusdensifloraSieb. & Zucc., 1842)[6]和我国广东珠海的黑松(PinusthunbergiiParl.)[7]上有分布,此次为我国华北地区的首次报道.本文对该北京油松线虫群体的形态和分子生物学特征进行描述,以期为今后该线虫的鉴定提供依据.
1 材料与方法
1.1 线虫分离
带有线虫的树枝样品采集于北京市郊的油松上,将其带回实验室,用斧头劈成细小片段.采用改良贝曼漏斗法[8]分离线虫,先向漏斗中注入适量自来水(以能恰好浸没样品为宜),于25 ℃左右静置24 h后,用凹面皿接取线虫悬浮液10 mL,在体视显微镜下检查.
1.2 形态学鉴定
将分离出来的雄虫和雌虫杀死固定后,采用甘油—乙醇快速脱水法脱水[9],然后挑入甘油滴中,用石蜡封片,制作成永久玻片.用Zeiss Imager Z1自动显微镜和Zeiss Axio Cam MRm数码相机观察形态并拍照,采用de Man公式对线虫进行测计[10].
1.3 分子生物学鉴定
单条线虫的DNA提取方法参见文献[11].rDNA 18S基因片段扩增引物及程序参照文献[12],rDNA 28S基因片段扩增引物及程序参照文献[13],rDNA ITS基因片段扩增引物及程序参照文献[14-15],扩增引物和退火温度见表1.将PCR产物分离纯化后,送上海美吉生物医药科技有限公司进行双向测序.

表1 本研究所用的引物序列和退火温度
用MEGA 6.0软件拼接测序所得序列,将拼接后的序列提交至GenBank获得登录号,并从GenBank下载相关线虫的序列进行比对分析,然后用MEGA软件构建Neighbor-joining系统进化树.
2 结果与分析
2.1 北京油松线虫群体的形态特征
从北京油松中分离的伞滑刃线虫群体雄虫和雌虫的形态测计值见表2.
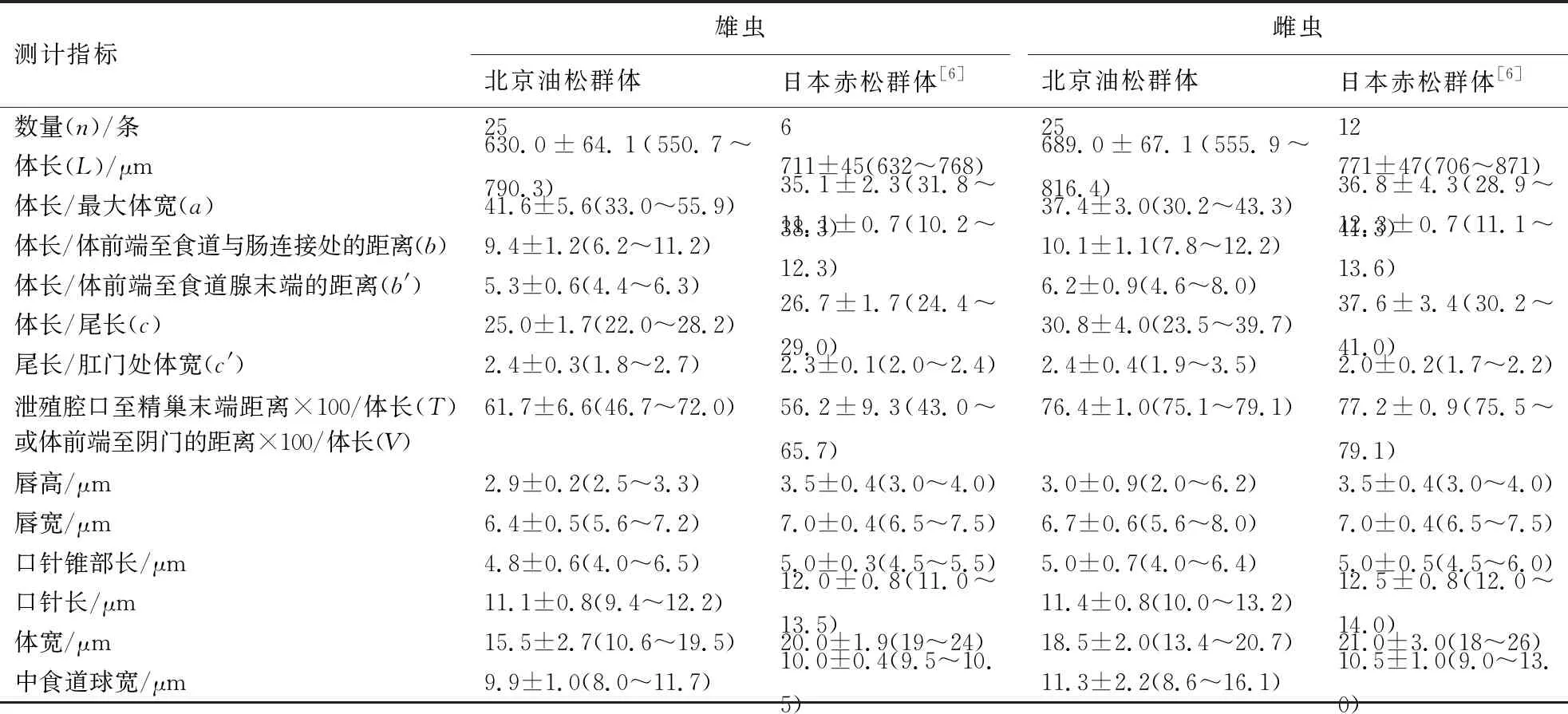
表2 东京伞滑刃线虫北京油松群体与原始文献形态测计值的比较
续表2
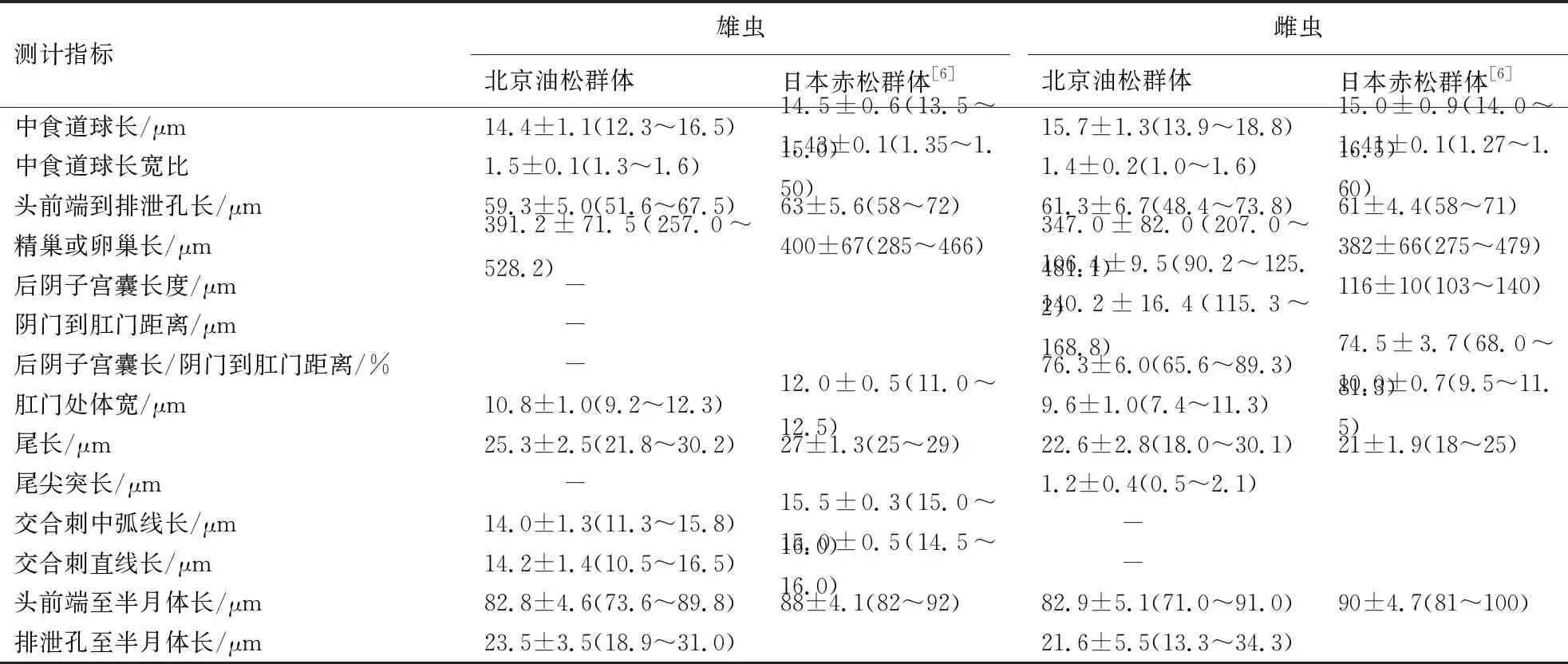
测计指标雄虫北京油松群体日本赤松群体[6]雌虫北京油松群体日本赤松群体[6]中食道球长/μm14.4±1.1(12.3~16.5)14.5±0.6(13.5~15.0)15.7±1.3(13.9~18.8)15.0±0.9(14.0~16.5)中食道球长宽比1.5±0.1(1.3~1.6)1.43±0.1(1.35~1.50)1.4±0.2(1.0~1.6)1.41±0.1(1.27~1.60)头前端到排泄孔长/μm59.3±5.0(51.6~67.5)63±5.6(58~72)61.3±6.7(48.4~73.8)61±4.4(58~71)精巢或卵巢长/μm391.2±71.5(257.0~528.2)400±67(285~466)347.0±82.0(207.0~481.1)382±66(275~479)后阴子宫囊长度/μm-106.4±9.5(90.2~125.2)116±10(103~140)阴门到肛门距离/μm-140.2±16.4(115.3~168.8)后阴子宫囊长/阴门到肛门距离/%-76.3±6.0(65.6~89.3)74.5±3.7(68.0~81.3)肛门处体宽/μm10.8±1.0(9.2~12.3)12.0±0.5(11.0~12.5)9.6±1.0(7.4~11.3)10.0±0.7(9.5~11.5)尾长/μm25.3±2.5(21.8~30.2)27±1.3(25~29)22.6±2.8(18.0~30.1)21±1.9(18~25)尾尖突长/μm-1.2±0.4(0.5~2.1)交合刺中弧线长/μm14.0±1.3(11.3~15.8)15.5±0.3(15.0~16.0)-交合刺直线长/μm14.2±1.4(10.5~16.5)15.0±0.5(14.5~16.0)-头前端至半月体长/μm82.8±4.6(73.6~89.8)88±4.1(82~92)82.9±5.1(71.0~91.0)90±4.7(81~100)排泄孔至半月体长/μm23.5±3.5(18.9~31.0)21.6±5.5(13.3~34.3)
雄虫:虫体圆柱形、细长,体前部渐细,热杀死后虫体向腹面弯曲呈“J”形(图1A),体表环纹明显,侧区有4条侧线(图1I).唇区缢缩明显,唇宽约为唇高的2倍(图1C).口针纤细,口针长9.4~12.2 μm,针锥长4.0~6.5 μm,约占口针总长度的44%,口针基部略增厚(图1C).中食道球卵圆形,后食道腺长,从背侧覆盖肠(图1B).排泄孔位置多变,一般位于中食道球对应处至中食道球后1个中食道球长度之间,半月体位于排泄孔后13.3~34.3 μm,一般位于神经环后.单精巢前伸,精巢有时回折;精母细胞在精巢前端1/3处多行排列,在精巢中部单行排列并形成大的精子,后部精子多行排列直至精巢末端.尾乳突7个:单个乳突(P1)恰好位于泄殖腔前腹中部,第1对亚腹乳突(P2)位于泄殖腔前,较P1略靠后或几乎平行,第2对亚腹乳突(P3)和第3对腹部乳突(P4)彼此接近,与P2相距约一个肛门处体宽.交合刺呈爪状,基顶小、方形、不显著;喙突小、钝尖;冠状体平整,在中部略凹陷;交合刺内缘和外缘光滑,无刻痕,逐渐变细,末端钝或略方,无盘状突(图1E-G).尾部短,向腹部强烈弯曲,约为肛门处体宽的2倍(图1E-G).交合伞卵圆形,末端钝尖(图1H).
雌虫:热杀死后虫体向腹面弯曲,在阴门处弯曲更加明显.体前部、食道等结构与雄虫相似.单卵巢前伸;卵母细胞在卵巢前端1/3处单行排列,在卵巢中部呈2~3行排列,在卵巢后部发育成单行成熟的卵细胞(图2A).阴门处具有一小阴门盖(图2B);后阴子宫囊很长,为阴门处体宽的6~8倍,约为肛阴距的76.3%,有时充满精子(图2A).肛门小,腹面观为一横裂.尾部短、近圆柱形,末端较宽,具有或尖或钝的短尾尖突,长0.5~2.1 μm(图2C-G).
东京伞滑刃线虫北京油松群体形态特征与Kanzaki et al[6]描述的日本赤松群体基本吻合.
2.2 与近似种的比较
根据4条侧线,雄虫交合刺似爪状、基顶不明显、喙突钝尖、无盘状突,雄虫尾突排列及数量,雌虫具小阴门盖、尾短、末端宽圆且有或钝或尖的尾尖突等特征,发现东京伞滑刃线虫北京油松群体与云杉伞滑刃线虫(B.piceaeTomalak & Pomorski, 2015)[16]、椴树伞滑刃线虫(B.tiliaeTomalak & Malewski, 2014)[17]、欧洲山毛榉伞滑刃线虫(B.fagiTomalak & Filipiak, 2014)[18]、指小伞滑刃线虫(B.idiusWalter Rühm, 1956)[19]十分相似.
东京伞滑刃线虫与云杉伞滑刃线虫的主要区别是交合刺形态,前者逐渐变细,而后者交合刺整体较粗,至交合刺末端才变细[16].
东京伞滑刃线虫与椴树伞滑刃线虫的主要区别:前者交合刺无盘状突,而后者有盘状突;交合刺中弧线长14.0(11.3~15.8) μm vs 20.0(18.1~21.8) μm;雄虫肛门处体宽10.8(9.2~12.3) μm vs 6.5(6.2~7.0) μm[17].
东京伞滑刃线虫与欧洲山毛榉伞滑刃线虫的主要区别:前者排泄孔位于中食道球对应处至中食道球后1个中食道球长度之间,后者位于中食道球对应处或略靠前; 前者交合刺逐渐变细,中部略粗,而后者急剧变细,中部略细;雄虫体长/最大体宽的值为41.6(33.0~55.9)vs 64.5(56.5~73.8),雌虫体长/最大体宽的值为37.4(30.2~43.3)vs 56.8(50.4~67.1)[18].
A.雄虫整体;B-D.体前部;E-G.尾部及交合刺;H.交合伞;I.侧线区.P1-P4.尾乳突.比例尺=10 μm.
图1 东京伞滑刃线虫北京油松群体的雄虫形态
Fig.1 Morphological characteristics of maleB.tokyoensis
A.后阴子宫囊;B.阴道、阴门;C-G.尾部.比例尺=10 μm.
图2 东京伞滑刃线虫北京油松群体的雌虫形态
Fig.2 Morphological characteristics of femaleB.tokyoensis
东京伞滑刃线虫与指小伞滑刃线虫的区别在于侧线数(4条vs 6条)和尾乳突位置(P2位于P1后,P3和P4靠近vs P2位于P1前,P3和P4分开)[19].
2.3 分子生物学特征
通过对北京油松线虫群体的rDNA扩增测序,共获得4条完全一致的长1 717 bp的18S序列,GenBank登录号为MK116557-MK116560,与日本赤松群体(登录号:AB430445)的相似度为99%.获得2条完全一致的长752 bp的28S-D2/D3序列,GenBank登录号分别为MK116561、MK116562,与日本赤松群体(登录号:AB430446)的相似度为99%.获得4条长948 bp的ITS序列,GenBank登录号为MK116563-MK116566,其中MK116563与MK116565的序列完全一致,且与MK116564序列有一个碱基的差异,与MK116566序列有4个碱基的差异,与日本赤松群体(登录号:AB430445)和广东珠海黑松群体(登录号:KY646465)的相似度分别为98%和97%.
基于rDNA 18S、28S和ITS基因序列构建了系统进化树(图3),发现北京油松群体均与日本赤松群体聚在一个分支上,相比与广东珠海黑松群体,北京油松群体与日本赤松群体的亲缘关系更接近.
综合形态特征和分子生物学特征,最终将该北京油松线虫群体鉴定为东京伞滑刃线虫.
3 讨论
结合文献报道[6-7]及本次调查可知,东京伞滑刃线虫的寄主比较广泛,赤松、黑松、油松均能被其侵染.
北京油松群体、日本赤松群体和广东珠海黑松群体形态特征基本一致,北京群体和日本群体测计值基本吻合,但是与广东群体存在较大的差异.广东群体雄虫体长/尾长的值为28.7~42.8,大于日本群体(24.4~29.0)和北京群体(22.0~28.2);泄殖腔口至精巢末端距离×100/体长的值为28.4~48.9,小于日本群体(43.0~65.7)和北京群体(46.7~72.0);肛宽5.8~8.8 μm,明显小于日本群体(11.0~12.5 μm)和北京群体(9.2~12.3 μm);精巢长177.5~282.1 μm,明显小于日本群体(285~466 μm)和北京群体(257.0~528.2 μm);尾长12.6~19.8 μm,明显小于日本群体(25~29 μm)和北京群体(21.8~30.2 μm).广东群体雌虫体前端至阴门的距离×100/体长的值为29.5~43.5,明显小于日本群体(75.5~79.1)和北京群体(75.1~79.1).此外,北京群体体长和体宽均略小于日本群体,这可能与日本群体测计的样本量较小或者地域不同有关系.
本研究在GenBank中提交的4条rDNA-ITS序列分别来自于同一寄主上的4条东京伞滑刃线虫,但这4条序列并不完全一致,说明来源于同一地域、同一寄主的东京伞滑刃线虫群体的rDNA-ITS序列存在种内变异,这为该线虫的鉴定增加了难度.
根据4条侧线,雄虫交合刺似爪状、基顶不明显、喙突钝尖,雌虫具小阴门盖、后阴子宫囊长约为肛阴距的3/4等特征,建议将东京伞滑刃线虫和云杉伞滑刃线虫、椴树伞滑刃线虫、欧洲山毛榉伞滑刃线虫、指小伞滑刃线虫归为一个组,称东京伞滑刃线虫组.基于ITS基因序列构建的系统进化树也支持该分组结论.另外,东京伞滑刃线虫日本赤松群体、广东珠海黑松群体和本研究中北京油松群体都是从枯死的松树上分离获得,对于此线虫能否致病,是否是导致松树死亡的原因,仍有待进一步研究.
