四川卧龙岷江冷杉天然林生物多样性随海拔梯度的响应
2020-07-09李旭华张临萍何飞冯秋红潘红丽蔡蕾宿以明刘兴良
李旭华, 张临萍, 何飞, 冯秋红, 潘红丽, 蔡蕾, 宿以明, 刘兴良*
1. 四川省林业科学研究院,四川 成都 610081;
2. 四川省林业和草原调查规划院,四川 成都 610081;
3. 四川省工程咨询研究院,四川 成都 610012;
4. 森林和湿地生态恢复与保育四川省重点实验室,四川 成都 610081;
5. 四川卧龙森林生态系统国家定位观测研究站,四川 阿坝 623006
植物群落多样性研究是群落生态研究中重要的内容[1]。植物群落多样性是指群落中物种的数目和每一物种的个体数目及其均匀度[2]。植物群落多样性随环境因子及演替梯度的变化特征是揭示生物多样性与生态因子相互关系的重要方面[3]。植物群落的梯度特征是指在群落组织水平上物种多样性的大小随某一生态因子梯度的有规律性的变化。山地植被群落生物多样性随海拔高度的变化规律一直是生态学家感兴趣的问题,海拔高度被认为是影响物种多样性格局的决定性因素之一[4-6]。
岷江冷杉(Abies faxoniana)林是我国特有的山地寒温性暗针叶林,特产于横断山区东北部[7],是四川省面积最大的原始森林类型和重要的用材林[8],也是长江和黄河上游重要的生态屏障[9]和大熊猫等珍稀野生动物的重要栖息地[10]。岷江冷杉种群在亚高山阴坡、半阴坡分布的海拔跨度特别大,同一山体坡面上海拔跨度可达1 200 m[11],环境梯度明显,各海拔岷江冷杉林内植被类型间的物种组成较丰富,层次结构明显。其林下分布着成片的华西箭竹(Fargesia nitida)、冷箭竹(Bashania fangiana)和峨眉玉山竹(Yushania niitakayamensis)等,气候温暖湿润,竹源丰富,是大熊猫的重要栖息环境。以往对岷江冷杉林的研究工作大多是关于群落特征、林窗特征等,对岷江冷杉林进行沿海拔梯度的生物多样性研究未见报道,因此,本研究试图通过对岷江冷杉林不同海拔生物多样性和物种相异性的测度,阐明物种沿环境梯度渐变或突变趋势以及植物群落在不同植被类型之内或之间的变化过程,探讨群落间的相似性程度和沿海拔梯度的分异规律,以期揭示岷江冷杉林群落多样性随海拔的变化规律,为川西亚高山暗针叶林天然更新、近自然人工林重建和生态系统管理等提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于四川省阿坝藏族羌族自治州汶川县境内的卧龙国家级自然保护区(30°45′—31°25′ N,102°52′—103°25′ E),地处青藏高原东南缘,川西高原向成都平原的过渡地带,属于四川盆地西缘山地[12]。该区域属典型亚热带季风气候,干湿季明显,据四川卧龙森林生态站(地点:汶川县卧龙镇邓生沟,海拔:2 700 m)观测资料,年平均温度8.4 ℃,1 月平均气温-1.7 ℃,7 月平均气温17.0 ℃,≥0 ℃活动积温4 000 ℃·d;年平均降水量861. 8 mm,5 月—9 月降水量占全年降水量的68.1%,年平均相对湿度80%左右[13]。全区地势起伏较大,由西北向东南递减,海拔范围1 150~6 250 m。植被类型丰富,随海拔梯度的变化形成明显的垂直分布格局,从沟谷到山顶依次分布着常绿阔叶林、落叶阔叶林、针阔混交林、亚高山针叶林、灌丛草甸等[14],其中,岷江冷杉暗针叶林在本研究区主要分布于海拔2 500~3 800 m 的山体阴坡上[11],土壤类型多样,形成与植被相适应的土壤结构与质地特征,主要有山地黄壤、山地黄棕壤、山地棕壤、山地棕色暗针叶林土和高山草甸土等[15]。
1.2 研究方法
1.2.1 样地调查与数据采集
在卧龙自然保护区邓生阴坡天然岷江冷杉林内沿海拔梯度2 700~3 600 m,每垂直上升200 m 设置一条调查样线,每条样线设置3 个样地,5 个海拔梯度共设置样地15 个,各个海拔梯度的基本情况如表1所示。采用分层取样的方法对岷江冷杉群落进行样地调查,每个样地内布设20 m×20 m 的乔木样方,在样方四角和中心设置5 个2 m×2 m 的灌木层样方和5 个1 m×1 m 的草本层样方,共得到乔木样方15 个,灌木样方75 个和草本样方75 个。调查并记录各样方的环境信息(包括经度、纬度、海拔、坡向、坡度、林冠盖度、病虫害和人为干扰状况等),乔木层所有胸径≥5 cm 的植物种类、每木胸径、树高、冠幅、枝下高、郁闭度、盖度等;灌木层植物种类、地径、高度、盖度及其数量;草本层植物种类、高度、盖度及其数量。
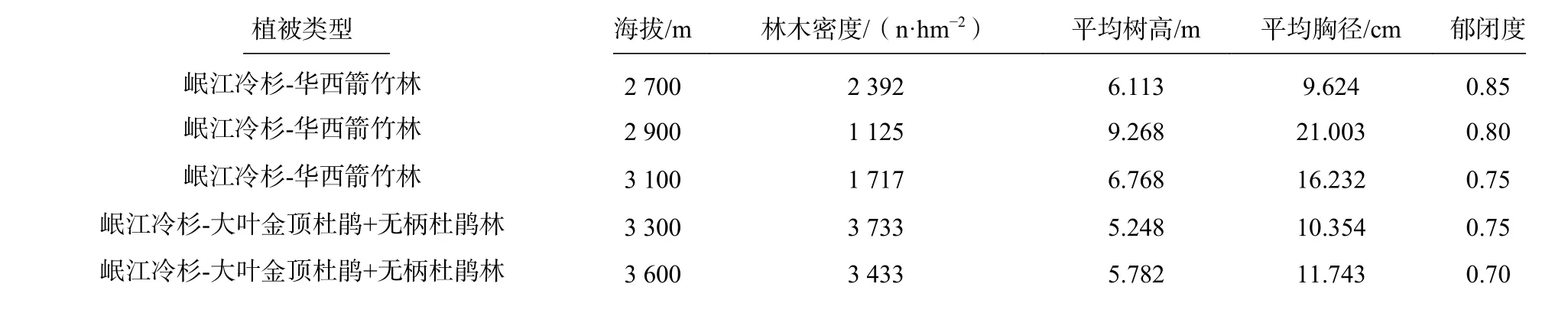
表1 不同海拔梯度岷江冷杉天然林的群落特征Tab. 1 Community characteristics of Abies faxoniana natural forest at different altitudes
1.2.2 数据分析与计算
1.2.2.1 物种重要值(IV)

1.2.2.2 α 多样性指数
α 多样性反映的是一个区域或一个生态系统可测定的生物学特征,它包括物种的种类和数量的丰富程度两方面[16]。本文采用以下5 个普遍应用的α 多样性指数来计算和分析各层的物种多样性和群落多样性[17]:



其中,S 为样方中记录到的物种数,Pi为i 物种的相对重要值,Ni为种i 的重要值,N 为S 个种的重要值之和。
1.2.2.3 β 多样性指数
β 多样性的测度主要有两种方式:基于物种组成的群落相异性(community dissimilarity)和基于物种分布界限的物种更替(turnover)。本文选取以下2 个指数对群落的β 多样性进行计算分析[18]:
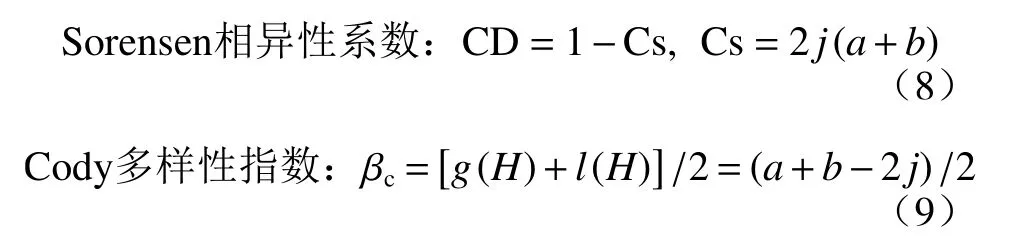
其中,g(H)为沿海拔梯度H 增加的物种数,l(H)为沿海拔梯度H 减少的物种数,j 为两群落中的共有物种数;a 和b 分别为该2 个群落中包含的物种数。
以上数据分析分别在Excel 和SPSS 软件中进行分析处理。
2 研究结果
2.1 不同海拔岷江冷杉天然林群落物种组成
卧龙岷江冷杉天然林5 个海拔上共调查到维管植物116 种,隶属42 科73 属。其中物种数目较多的有8 个科,共27 属56 科(见表2),分别占样方内所有物种属、种的64.29%和48.28%。蔷薇科和槭树科为乔木层常见种;杜鹃花科为高海拔处灌木层优势种,蔷薇科、忍冬科和卫矛科为灌木层常见种;菊科多为草本层优势种,百合科和毛茛科为草本层常见种。

表2 5 个海拔岷江冷杉天然林优势科的属、种统计Tab. 2 Statistics of the genus and species of dominant families of Abies faxoniana natural forest at five altitudes
2.2 不同海拔岷江冷杉天然林群落α 多样性
2.2.1 不同海拔群落乔木层的α 多样性
乔木种在5 个海拔上的α 多样性分析结果表明:随着海拔梯度的上升,卧龙岷江冷杉林的乔木物种多样性下降。其中,乔木层的物种丰富度、Shannon-Wiener 多样性指数和Pielou 均匀度指数变化一致,均呈下降趋势,而生态优势度指数则呈上升趋势(见表3),均优丰指数计算结果与Shannon-Wiener 多样性指数相一致。

表3 岷江冷杉林乔木层α 多样性指数在不同海拔梯度上的变化Tab. 3 Variations of tree species α diversity index in Abies faxoniana forest at different altitudes
2.2.2 不同海拔群落灌、草层的α 多样性
卧龙岷江冷杉林5 个海拔上的灌木种随着海拔梯度上升物种丰富度变化与乔木种呈相同的下降趋势。而Shannon-Wiener 多样性指数、Pielou 均匀度指数和均优丰指数则呈现出先降低后升高的趋势(见表4),在海拔2 900 m 处最低,而生态优势度指数的变化则与之相反,呈现出先增加后减小的趋势,2 900 m 处的优势度指数最大,为0.236。
草本植物的α 多样性指数变化与乔木层基本相同,即随着海拔梯度上升草本物种丰富度、Shannon-Wiener 多样性指数、Pielou 均匀度指数和均优丰指数均呈下降趋势,而生态优势度指数呈上升趋势(见表5)。与乔灌层相比,草本层的多样性和均匀度较高,且随海拔的变化幅度也较小。

表4 岷江冷杉林灌木层α 多样性指数在不同海拔梯度上的变化Tab. 4 Variations of shrub species α diversity index in Abies faxoniana forest at different altitudes

表5 岷江冷杉林草本层α 多样性指数在不同海拔梯度上的变化Tab. 5 Variations of herbaceous species α diversity index in Abies faxoniana forest at different altitudes
2.2.3 不同海拔群落α 多样性
综合分析群落的多样性,结果显示(见表6):随着海拔梯度的上升群落所有植物的物种丰富度指数、Shannon-Wiener 多样性指数、Pielou 均匀度指数和均优丰指数均呈下降趋势,而生态优势度指数呈上升趋势。卧龙岷江冷杉林群落在低海拔处的生物多样性最高,优势度最低,稳定性最好。随着海拔的上升,水、热、光等生态因子发生相应改变,生物多样性明显减小,优势度升高,群落稳定性降低。

表6 岷江冷杉林群落的α 多样性指数在不同海拔梯度上的变化Tab. 6 Variations of community species α diversity index in Abies faxoniana forest at different altitudes
2.3 不同海拔岷江冷杉天然林群落β 多样性
2.3.1 不同海拔群落乔木层的β 多样性
通过对乔木层物种的β 多样性测度,结果表明:随着海拔上升,相邻海拔间的相异系数CD 和Cody 指数呈下降趋势,即海拔越高,相邻海拔间的乔木物种相似性越高,更替速率越慢(见图1)。
2.3.2 不同海拔群落灌、草层的β 多样性
灌木物种的β 多样性分析表明:随海拔上升,相异系数CD 和Cody 指数没有出现统一的上升或下降的趋势,而是在海拔2 700~2 900 m 和海拔3 100~3 300 m 两个海拔间形成双峰值。由于海拔2 700~2 900 m 物种数目急剧减少,所以导致了β 多样性较高;而海拔3 100~3 300 m 由于灌木层的优势种发生改变,由华西箭竹变成了杜鹃,因此β 多样性也相对较高(见图2)。
草本植物的β 多样性变化情况与乔木一致,相异系数CD 和Cody 指数随海拔的升高而降低(见图3),即随着海拔的升高,岷江冷杉林相邻海拔间的草本植物相似度越高,但草本层的Cody 指数较乔木层和灌木层高。

图1 乔木层β 多样性在不同海拔梯度上的变化Fig. 1 Variations of tree layer β diversity in Abies faxoniana forest at different altitudes
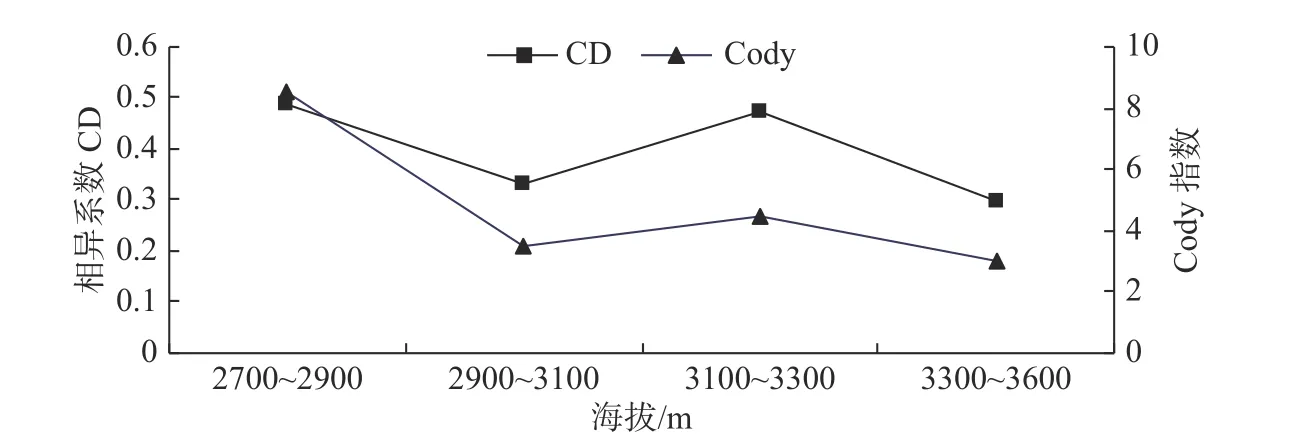
图2 灌木层β 多样性在不同海拔梯度上的变化Fig. 2 Variations of shrub layer β diversity in Abies faxoniana forest at different altitudes
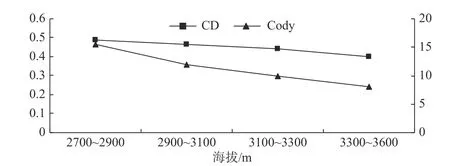
图3 草本层β 多样性在不同海拔梯度上的变化Fig. 3 Variations of herbaceous layer β diversity in Abies faxoniana forest at different altitudes
2.3.3 不同海拔群落β 多样性

图4 岷江冷杉林群落β 多样性在不同海拔梯度上的变化Fig. 4 Variations of β diversity in Abies faxoniana forest community at different altitudes
综合分析群落总的β 多样性发现,相异系数CD 和Cody 指数均随海拔的升高而明显降低,后者反应更明显,这与乔木和草本的变化一致,而与灌木的变化不同(见图4);而群落总的Cody 指数变化主要受草本层影响。由于乔木和草本的物种数在群落中占大多数,因此群落物种β 多样性的变化与之相同,可见群落β 多样性的变化主要由物种数大的植被类型所决定。对比各生活型植物的β 多样性可见:各海拔群落相异系数(CD)表现出乔木>群落>草本>灌木的规律;而Cody 指数都具有群落>草本>乔木>灌木的规律。
3 讨论
3.1 岷江冷杉林α 多样性的海拔梯度格局
卧龙岷江冷杉天然林物种α 多样性随海拔梯度的垂直变化表明:不同生长型植物的物种α 多样性随海拔梯度的变化趋势各有不同。乔木和草本植物随着海拔梯度上升物种丰富度、Shannon-Wiener 多样性指数、Pielou 均匀度指数和均优丰指数均呈下降趋势,其中均优丰指数的计算使不同群落多样性指数对比差异更加明显,且能够突出物种数在群落多样性中的重要作用[19]。而生态优势度指数呈上升趋势。对于木本植物,其物种多样性随海拔升高而降低的变化规律较为普遍地存在于不同的生态系统[20-21],但也有研究表明木本植物多样性随海拔呈明显的单峰型变化[22]。而草本植物多样性随海拔变化的关系较为复杂,其随海拔变化而下降的变化规律常见于温带森林[20]和热带森林中[23]。
灌木种则随着海拔梯度上升物种丰富度下降;而Shannon-Wiener 多样性指数、Pielou 均匀度指数和均优丰指数则呈现出先降低后升高的趋势;生态优势度指数的变化与之相反。这是由于物种丰富度只反映了物种在群落中的存在与否,而Shannon-Wiener 多样性指数、Pielou 均匀度指数和均优丰指数还加入了物种数量多少的影响因素。调查中发现,在海拔2 700 m 处虽有华西箭竹生长,但由于该海拔物种丰富,在多物种的竞争下华西箭竹的优势度并未特别突出;而在海拔2 900 m 和3 100 m 处,物种丰富度下降明显,竞争减小,华西箭竹这一个种占据了灌木层明显的主导地位,其优势度远远大于其他灌木种,从而大大降低了该群落的均匀度,造成了这两个海拔上的灌木多样性较低;到了高海拔3 300 m 以上时,没有了华西箭竹的分布,杜鹃占据灌木层优势地位,但其优势度与其他物种没有太大差距,所以高海拔处生物多样性上升。
群落所有植物的α 多样性表明:随着海拔梯度的上升群落的物种丰富度、Shannon-Wiener 多样性指数、Pielou 均匀度指数和均优丰指数均呈下降趋势,而生态优势度指数则与其他几个指数呈负相关,它的值越高说明群落中仅有少数优势种,群落不稳定[24]。海拔变化是决定本研究区生境差异的主导因子,海拔导致水热条件及其组合的空间分布,进而影响植物群落的分布及结构。迄今为止,有关山地森林群落物种多样性沿海拔梯度的变化模式虽然尚无统一认识,但诸多研究表明,物种多样性随海拔升高而逐渐减少是山地物种多样性垂直格局变化的一种趋势[25-26]。此次研究结果也显示卧龙岷江冷杉林群落的生物多样性随海拔上升呈下降的趋势,与其他研究的结果一致。
3.2 岷江冷杉林β 多样性的海拔梯度格局
β 多样性可以定义为沿着环境梯度的变化物种替代的程度,亦有学者称之为物种周转速率、物种替代速率和生物变化速率[27-28]。它能够比较直观地反映不同群落间物种组成的差异[29],可用于分析不同生境间的梯度变化,呈现群落间的相互关系以及物种沿着环境梯度的替代规律[30-32]。通过对群落β 多样性的测度,可以揭示生境被物种分割的程度或不同地段的生境多样性[33]。多数研究表明,植物的β 多样性随海拔升高而单调下降[23,34]。本文研究结果也表现出同样的变化规律,即随着海拔上升,岷江冷杉林内乔木、草本和所有群落所有植物的相异系数CD 和Cody 指数均呈下降趋势;这种β 多样性在低海拔地区较高的现象可能是由于干扰和物种间的相互竞争,中度干扰在增加了群落α 多样性的同时,也导致了生境的异质性,从而导致β 多样性的增加[34]。而灌木层因为物种数和优势种的改变形成β 多样性的双峰变化,这与灌木植物的实际分布是相符的。整体群落的物种差异和更替速率受物种数较多的植被类型影响较大,随海拔升高呈逐渐减小趋势。形成这种变化的原因可能是因为卧龙岷江冷杉林随着海拔的升高,物种数明显下降,减少的物种数导致了更低的物种更替速率,因此物种数是影响β 多样性的重要因素。
