稻米镉积累特性及其遗传调控研究进展
2020-07-07李炜星欧阳林娟陈小荣彭小松贺浩华孙晓棠朱昌兰
李炜星 欧阳林娟 王 薇 陈小荣 彭小松 贺浩华 孙晓棠 朱昌兰
(江西农业大学作物生理生态与遗传育种教育部重点实验室,南昌 330045)
稻米是我国三分之二以上人口的主食,其质量安全直接关系到人民健康。近年来,工业“三废”排放、污水灌溉、含镉投入品使用、土壤酸化促进有效镉转化等,导致部分农田镉含量超标。镉污染不仅影响水稻生长发育,使产量下降,更严重的是会导致镉在稻米中超标累积,危及稻米安全。镉被世界卫生组织划为一级致癌物质,在人体内的半衰期为10~30年,长期食用镉超标食品将导致慢性中毒,引发骨质疏松、肾功能衰竭、癌症及心血管疾病等。近年来,稻米镉含量超标现象在我国时常发生,已成为影响人类健康的重大安全问题。利用作物本身对镉积累的差异,选育籽粒低积累的水稻品种,是防控治理稻米镉污染问题最直接和经济有效的途径。本文从不同基因型水稻籽粒镉积累差异、稻米中镉的分布及存在形态等方面综述了稻米镉积累特性,并从镉积累相关基因及数量性状基因座(quantitative trait locus;QTL)的鉴定与功能分析等方面综述了稻米镉积累遗传调控研究进展,对稻米镉积累的研究前景进行了展望,旨在为降低稻米镉含量、减少食品安全问题提供借鉴。
1 稻米镉积累特性
许多研究表明,镉在稻米中的积累存在显著的基因型差异,这为通过遗传改良降低稻米镉含量提供了可能。多数研究结果认为,籼稻吸收和转运镉的能力大于粳稻[1-5]。有研究发现,杂交稻比常规稻更容易吸收和富集镉[6],与普通杂交稻相比,超级杂交稻籽粒的Cd含量较高,锌含量较低[7]。也有研究认为杂交籼稻与常规籼稻品种相比,籽粒对镉的累积能力不存在明显差异,不能按水稻类型来筛选镉低积累品种[8,9]。由于晚稻生长季节根际土壤镉含量显著高于早稻生长季节,晚稻的稻米镉超标情况往往较早稻更为严重[10]。
镉在稻谷中的分布不均匀,主要分布在水稻的胚、糊粉层和外层胚乳中,胚乳中分布较少[11]。水稻各部分镉含量由高到低为皮层>胚>胚乳>颖壳[12],也有认为颖壳的镉含量高于胚乳,胚乳中的镉含量最低[13]。因此,稻谷经加工后,米糠中富集了很大一部分镉,其镉浓度可比精米高5倍以上[2]。虽然米糠只占稻谷总质量的9%,精米占稻谷总质量的70%,但米糠中富集的镉元素约占整个稻谷籽粒镉元素的40%。
镉在稻米中主要以蛋白结合态形式存在,占水稻籽粒镉总量53.4%,淀粉结合态占12.3%,其他形态占34.3%[11]。不同蛋白组分的镉结合能力表现为球蛋白镉浓度最高,清蛋白次之,醇溶性蛋白和谷蛋白镉浓度则相对较低,但球蛋白和谷蛋白中镉的总含量处于遥遥领先的地位[11]。清蛋白和球蛋白主要集中在米糠中,谷蛋白主要分布在胚乳中,醇溶性蛋白的分布比较均匀,不同部位含量差异不明显[11]。发现镉主要与54.5 ku和5.5 ku分子质量的蛋白结合,富镉蛋白中谷氨酸、缬氨酸、半胱氨酸、异亮氨酸、亮氨酸、酪氨酸的相对含量较高[14]。研究表明含硫氨基酸残基可能是蛋白质与镉结合的位点[11]。
2 稻米镉积累相关基因及其功能
镉在稻米中的积累主要是通过根系对镉的吸收,经过木质部的装载和运输转移到地上部,再经过韧皮部的运输和转移积累到籽粒中。现有研究表明,自然抗性巨噬细胞蛋白(natural resistance-associated macrophage protein,NRAMP)、重金属 ATP 酶(heavy metal ATPase, HMA)、锌铁转运蛋白(ZIP)家族等一些转运蛋白基因参与了水稻对镉的吸收和积累(表1)。
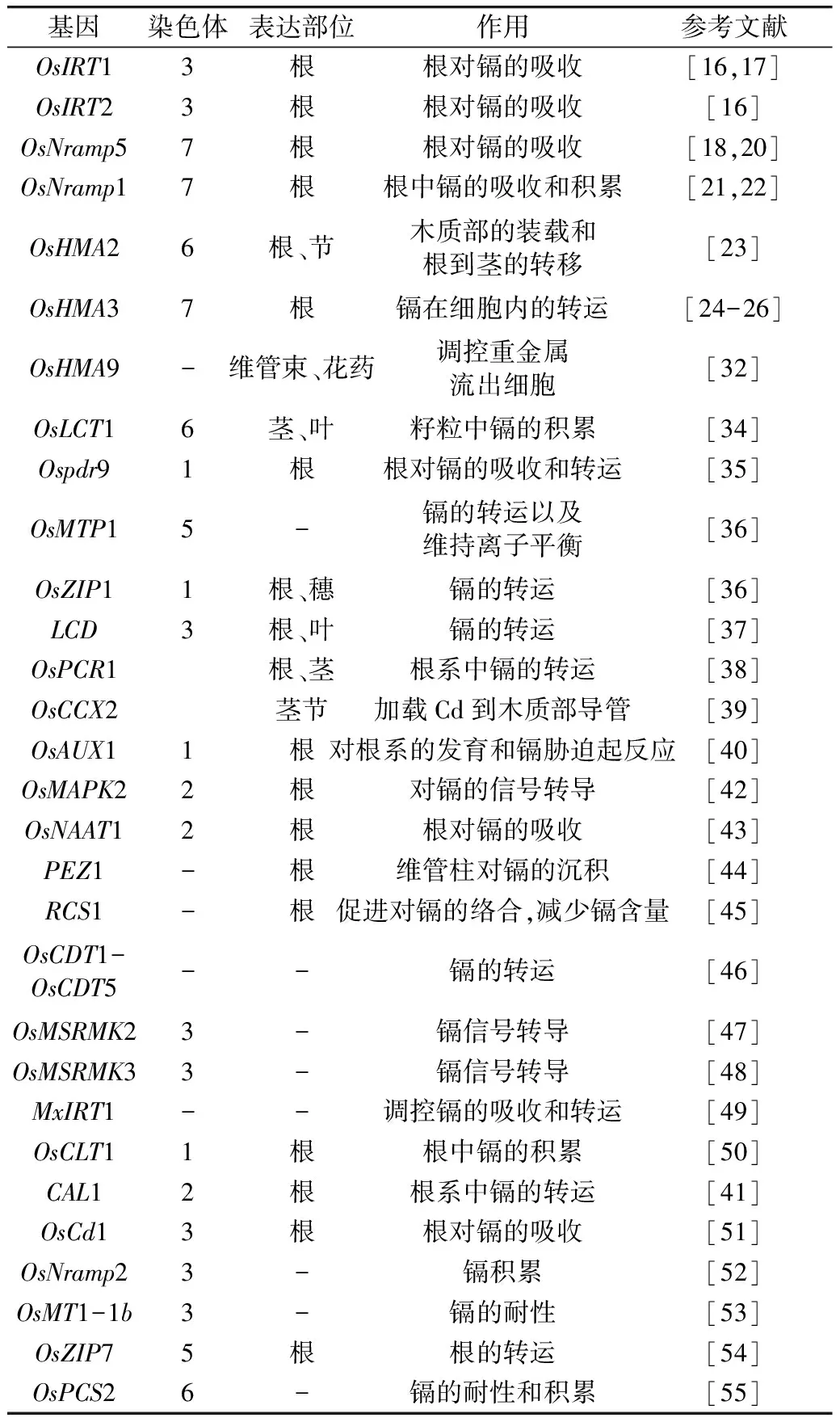
表1 已鉴定的稻米镉积累相关基因
2.1 锌铁转运蛋白基因
Pence等[15]首先发现镉离子可以通过水稻锌铁转运蛋白在水稻体内进行转运。Nakanishi等和Lee等分别报道了Zn2+和 Fe2+的转运蛋白OsIRT、OslRT2对 Cd2+也具有较高的亲和性,在缺铁处理下能增强水稻对 Cd2+的吸收和转运[16,17]。
2.2 NRAMP家族基因
NRAMP是一类具有二价金属离子转运功能的蛋白,能够转运Mn2+、Zn2+、Cu2+、Fe2+、Cd2+、Ni2+、Co2+和Al3+。水稻基因组中有7个OsNRAMP,其中OsNramp5和OsNramp1 已被证实参与了调控Cd的吸收与运输。OsNramp5能够调控Mn2+、Fe2+和Cd2+等穿过凯氏带进入木质部维管束[18],缺失OsNramp5在降低镉积累的同时,也显著降低水稻生长和籽粒产量[19,20]。
与OsNramp5相比,OsNramp1基因的作用很小。OsNramp1蛋白参与镉、铁的吸收和转运[21]。OsNramp1主要在水稻根部表达,参与中柱和木质部中镉的装载转运。研究发现,OsNramp1在水稻品种Sasanishiki和Habataki的根系中表达水平差异明显,导致2个品种镉积累量差异显著[21]。有研究发现OsNramp1在粳稻中的表达水平明显低于籼稻[22]。
2.3 重金属 ATP 酶(HMA)基因
HMA是一种重金属离子跨膜蛋白,通过 ATP 水解提供的能量实现重金属离子在细胞膜上的跨膜转运。水稻全基因组中有9个编码HMA的基因(OsHMA1~OsHMA9),其中OsHMA1~OsHMA3属于 Zn/Co/Cd/Pb二价阳离子结合型,其余 6个属于 Cu/Ag 单价阳离子结合型[23]。Ueno等[24,25]和Miyadate等[26]团队分别通过图位克隆技术分离到位于第7染色体上的单隐性基因OsHMA3,这是水稻中迄今为止唯一一个通过正向遗传学手段克隆得到的Cd 相关基因。OsHMA3可以将由根吸收到的Cd2+封存在根部细胞的液泡中,从而降低由木质部向地上部运输的Cd2+量,导致籽粒的Cd积累减少[26,27]。过表达OsHMA3能显著降低Cd在水稻籽粒中的积累,而不影响Zn、Fe等其他金属离子的运输和积累。也有研究发现OsHMA3过表达还能提高根部中的锌浓度,上调茎秆中部分锌转运蛋白基因的表达量[28]。在高Cd积累的籼稻品种中发现了OsHMA3的等位突变[25,29],在粳稻中也发现了OsHMA3的等位突变,无论盆栽还是大田试验下,突变粳稻品种中籽粒的Cd 含量均显著高于普通粳稻品种[30]。
OsHMA2是OsHMA3的一个同源基因,主要在根系中表达,在茎节中有少量表达。与OsHMA3不同的是,OsHMA2 是一个外转运子,OsHMA2的过表达能加速镉、锌等金属离子向木质部的装载及其从地下部向地上部的转运,增加叶片中镉和锌的含量。与野生型相比,过表达OsHMA2能使籽粒Cd含量降低一半,而对Zn等其他元素含量没有影响;抑制OsHMA2表达则能降低叶片中的Cd、Zn 含量,提高根系中的Zn含量[23,31]。
OsHMA9主要在叶肉细胞和花药中表达,虽然属于Cu/Ag亚群,负责细胞中Cu、Zn、Pb的向外转运,但也与木质部Cd 的装载有关,可能是镉的另一个外转运子,参与细胞内镉等金属离子的向外转运[32,33]。高浓度的Cd可诱导OsHMA9在维管束中大量表达,OsHMA9缺失突变体中Cd积累量大大提高[34]。
2.4 其他转运蛋白基因
OsLCT1是镉低亲和性阳离子转运蛋白基因(Low-affinity cationtransporter,LCT),是一个韧皮部转运子,主要在水稻灌浆过程中将镉从茎节经韧皮部转运到籽粒中[35]。OsPDR9是一个ATP-结合框蛋白(ATP-binding cassette, ABC)基因,受Cd诱导在水稻幼苗根系大量表达,可能影响水稻根系中Cd的积累[36]。OsMTP1和QZT1同为金属耐受蛋白(metal tolerance protein, MTP)基因,编码阳离子扩散促进因子蛋白,通过高效转运Cd、Zn和其他重金属元素,维持植物体内阳离子的平衡,受 Cd2+、Zn2+、Mg2+、Cu2+和 Fe2+等重金属离子诱导[37]。低镉基因(low cadmium, LCD)在根维管束和叶片韧皮部伴胞中高表达,与野生型相比,突变体叶片的Cd 含量没有差异,谷粒的Cd含量减少了约50%,表明突变体运输和积累镉的能力降低,推测OsLCD可能是参与调控韧皮部中Cd 转运的基因[38]。镉抗性相关蛋白基因(plant cadmium resistance 1, PCR1)主要表达部位为幼苗期的根和生殖期的第Ⅰ节、第Ⅱ节间,在水稻穗中检测有一定表达量;OsPCR1敲除株系的籽粒表现出镉含量降低,但同时伴随着粒重下降和粒长变短的不良变化[39]。OsCCX2是一个在茎节表达的转运蛋白基因,可能是一个阳离子/钙(Ca)交换器,负责Cd加载到木质部导管中,从而介导镉在水稻籽粒中的积累[40]。CAL1功能缺失的突变体中水稻叶片中的镉浓度显著降低[41],其他转运蛋白基因还有OsAUX1、OsMAPK2、OsNAAT1、PEZ1、RCS1、OsCDT1-OsCDT5、OsMSRMK2、OsMSRMK3、MxIRT1、Os-ZTP6、OsFRDL1等[42-50]。
3 稻米镉积累相关QTL分析
Ishikawa 等[56]首次报道利用越光和Kasalath的CSSL群体在3、6、8 号染色体上定位到3 个控制糙米Cd 含量QTL,随后国内外学者利用各种遗传分离群体,通过连锁分析和全基因组关联分析,定位到了至少100余个稻米镉积累相关QTL,分布在水稻的12条染色体上(表2)[24,29,57-71]。
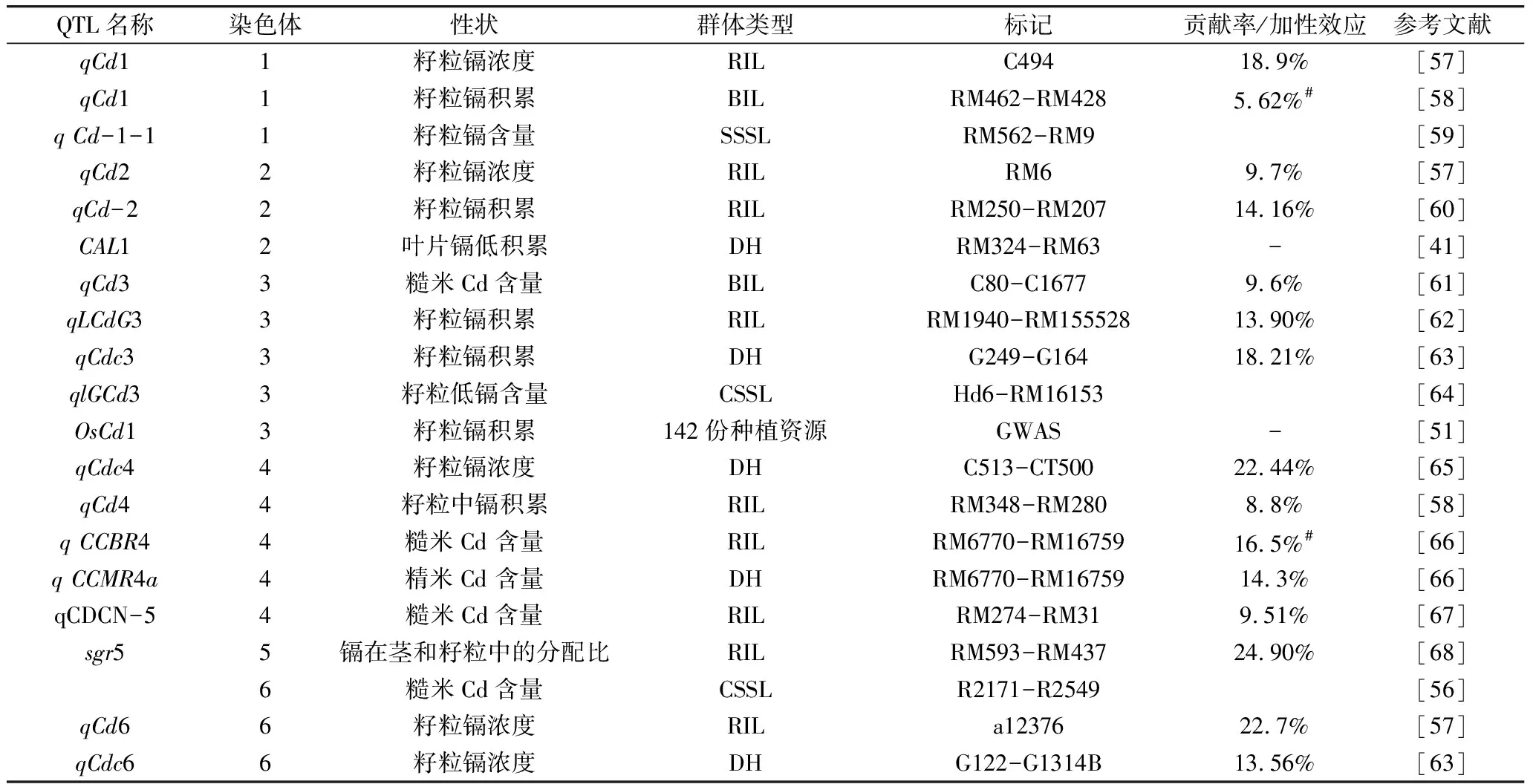
表2 稻米镉积累相关QTL

续表2
注: 重复定位的QTL未列入。*已经克隆;#平均值。
总体来看,除了在第7染色体短臂上检测到一个贡献率较大的QTL外,检测到的QTL效应均较小,贡献率基本在20%以下。虽然有多个QTL被不同学者定位在了相同的区域,但在不同群体和试验中重复定位到的QTL总体较少。目前已进行精细定位的QTL只有4个[24,29,50,51,61],通过进一步图位克隆并得到鉴定的QTL仅有3个。Ueno等[24]、Tezuka等[29]分别利用低镉品种和高镉品种的F2群体,均在第7染色体短臂上定位到一个效应很大的QTL,控制根部Cd向茎转移率,经过进一步精细定位,克隆了该QTL,验证发现与OsHMA3为同一个基因。LUO等[41]利用TN1和CJ06构建的双单倍体群体进行QTL定位分析, 克隆到1个特异调控镉在水稻叶片中积累的主效QTL基因CAL1。
4 展望
镉是一种严重危害人体健康的重金属元素。在分子水平上,研究者们已经鉴定出一些金属转运蛋白基因参与调控水稻对镉的吸收和积累,如NRAMP1、NRAMP5、HMA2、HMA3等,研究者通过遗传改良调控这些基因的表达,阻断镉的吸收、运转,获得了镉吸收和积累明显降低的水稻材料。如Ishikawa等[19]通过辐射获得OsNramp5缺陷突变体,大幅度降低了根部对 Cd2+的吸收,使得稻米镉含量降低到了安全阈值。借助蔗糖转运蛋白基因OsSUT1启动子调控OsHMA2特异表达使种子中的镉含量降低到野生型的一半[23]。湖南杂交水稻研究中心通过CRISPR/Cas9基因编辑技术定点突变OsRAMP5基因,培育出“低镉1号”等低镉水稻品种。但是,由于这些金属转运蛋白基因的底物广泛,能够同时运输多种必需营养元素(如Mn,Fe 和Zn),通过调控这些基因的表达,在降低水稻对Cd 吸收和积累的同时,也可能对必需矿质营养元素的吸收和积累产生影响。因此,在利用这类基因时,如何避免其降低稻米营养品质所产生的负效应是必须要考虑的问题。
目前关于水稻Cd吸收和积累的QTL定位研究有不少,但通过QTL定位并最终克隆鉴定的基因只有OsHMA3一个,精细定位的QTL也极少,究其原因可能是水稻对Cd的吸收和积累的遗传比较复杂,作为多基因控制的数量性状,QTL的效应值较小,不容易鉴别,且QTL的表型效应受环境影响大。利用染色体片段置换系或近等基因系,减少遗传背景干扰,尽量使QTL解离为单个孟德尔因子,将有助于QTL的精细定位和图位克隆。此外,利用转录组学、蛋白质组学、代谢组学等多组学结合的方法,通过高通量技术快速分离更多的镉胁迫响应基因,更加深入地解析其遗传调控网络,探究其在水稻生长过程中的作用,从而为降低稻米镉含量、减少食品安全问题提供参考。
