PRC1对于小鼠卵母细胞作用机制的初步研究
2020-07-07王新洁许钟峯王海龙
王新洁,许钟峯,王海龙
(厦门大学 医学院,福建 厦门361102)
哺乳动物的卵母细胞指雌性动物的生殖细胞,一般呈球体,且相较于体细胞体积偏大,它不仅具有一般体细胞共有的结构,而且含有受精后胚胎早期发育所需的各种营养物质。卵母细胞的发育成熟是通过减数分裂进行的,在这个过程中卵母细胞进行1次DNA复制和2次连续不均等分裂,形成染色体倍数减半的单倍体配子,且将大部分营养物质集中在成熟卵母细胞中,以保证卵母细胞正常受精和母源遗传物质的稳定[1]。
1 文献综述
减数分裂是卵母细胞发育成熟的重要基础。以小鼠卵母细胞成熟过程为例,哺乳动物的卵母细胞在到达性成熟期前一直停滞在分裂前期,此时细胞核呈现膨大的状态,核膜完整,染色质高度疏松,该时期的细胞核呈泡状,称为生发泡,这一时期又称为生发泡期(GV)。进入青春期后,卵母细胞受到促性腺激素的刺激恢复减数分裂,核膜破裂,染色质外流,生发泡破裂(GVBD,标志着减数分裂的恢复)。GVBD之后,染色质不断浓缩成染色体,微管蛋白开始在微管组织中心的调控下在其周围组装微管。随着微管不断组装,在卵母细胞中央大量微管纵向排列,组成中间宽、两极小的纺锤体,染色体移向赤道板,卵母细胞进入减数分裂前中期(pro-MⅠ)。之后纺锤体被以微丝为主的胞质流动牵引着向纺锤体长轴方向移动,逐渐迁移到卵母细胞皮质膜下方。此时在靠近纺锤体一端的细胞皮质中微丝开始活跃地组装,并在皮质层下方聚集呈现一个帽状分布,称为微丝帽,此时卵母细胞进入第一次减数分裂的中期(MⅠ)。MⅠ期之后,在双极纺锤体的拉动下,同源染色体开始分离,进入第一次减数分裂末期(TI)。此时纺锤体进一步拉动同源染色体向两极分离,纺锤体突出细胞边缘,接着一半的染色体从卵母细胞分离,排出体积比较小的第一极体(PBI)。PBI排出后,卵母细胞内第二次减数分裂的纺锤体迅速组装完成,微管牵引着卵母细胞内的染色体重新排列在赤道板上,卵母细胞进入第二次减数分裂的中期(MⅡ)。此时标志着卵母细胞成熟;成熟的卵母细胞停滞于MⅡ期,等待精子的到来,再继续完成第二次减数分裂的过程。减数分裂的失败可能会导致受精异常、早期胚胎发育阻滞和不良妊娠结局等。因此,充分掌握卵母细胞减数分裂调控机制对于提高卵母细胞质量、完善以卵母细胞为基础的辅助生殖技术是十分重要的。小鼠卵母细胞减数分裂的过程见图1。
PRC1是微管结合蛋白家族中的重要成员之一,是细胞周期蛋白依赖性激酶的底物。D.Pellman等[2]于1995年首次在酵母中发现了PRC1的同源物Ase1蛋白,并且证明其绑定于有丝分裂纺锤体微管,作用于有丝分裂后期纺锤体移动的过程。随后Jiang W.等[3]于1998年在HeLa细胞中发现并命名了PRC1蛋白,并且阐明PRC1在细胞有丝分裂的胞质分裂过程中发挥了关键性作用。接着Y.Kurasawa等[4]在2004年发现KIF4和PRC1绑定在一起,且对于有丝分裂纺锤体的形成起着至关重要的作用。另外一项研究表明,在有丝分裂后期PRC1可以识别纺锤体微管末端的重叠区域,阻止纺锤体的解聚[5]。在最近的研究中,Guo J.等[6]于2018年在卵母细胞中发现PRC1蛋白在雷帕霉素机制性靶标通路中起着重要作用,它对于减数分裂进行到MⅡ期是十分重要的,揭示了PRC1在生殖细胞成熟发育过程中发挥重要功能。
通过20余年的研究,学者们对PRC1的结构和功能有了一定的了解,PRC1在纺锤体的组装和胞质分裂中都发挥着重要作用。虽然目前的研究表明PRC1在卵母细胞中发挥了一定作用,但在哺乳动物卵母细胞减数分裂中PRC1发挥何种功能、具体的作用机制又该如何描述,均有待于进一步深入探究。因此本试验主要探讨PRC1在小鼠卵母细胞减数分裂中的作用,从而对PRC1在小鼠卵母细胞中的作用机制进行初步研究。
2 材料与方法
2.1 试验动物
7~8周龄雌性昆明小白鼠(体重22~28 g),购自上海斯莱克实验动物有限公司。小鼠饲养在厦门大学实验动物中心,环境为“无特定病原体”级别,温度控制在22℃左右。配制可供小鼠自由取食的标准鼠粮和无菌水。光照周期设置为12 h光照、12 h黑暗。
2.2 试剂与仪器
兔源抗PRC1抗体,购自艾博抗(上海)贸易有限公司;CyTM5偶联羊抗兔二抗,购自Jackson Immuno Research公司;防荧光猝灭的DAPI封片剂,购自Vector Laboratories公司;微量RNA提取试剂盒、逆转录试剂盒,均购自赛默飞世尔科技(中国)有限公司;qRT-PCR所用试剂套装,购自北京全式金生物技术有限公司;其他试剂如无特殊说明,均购自西格玛奥德里奇(上海)贸易有限公司。
高灵敏度激光共聚焦显微镜(型号为Zeiss LSM 880+Airyscan),购自卡尔蔡司公司;实时定量PCR仪(型号为CFX Connect Optics Module),购自伯乐公司;CO2培养箱(型号为3111),购自赛默飞公司;体式显微镜(型号为Nikon SMZ1000),购自尼康公司。
qRT-PCR引物由生工生物工程(上海)股份有限公司设计并合成,序列见表1。
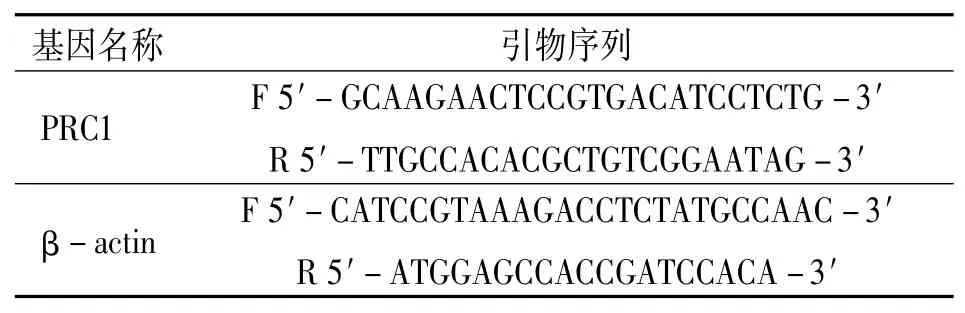
表1 用于qRT-PCR的基因引物序列
2.3 关键试验方法
2.3.1 卵母细胞的采集与培养 采用脊椎脱臼法处死小鼠,快速分离双侧卵巢。清洗干净后用无菌刀片将卵巢充分剁碎,而后加入适量M2培养液摇匀。将培养基置于体视显微镜下,利用装上拉制玻璃管(北京正天贸易有限公司)的口吸管挑选出形状正常、周围无颗粒细胞带有生发泡的卵母细胞,充分洗净后转移至覆盖石蜡油的M2培养液中,然后将培养皿放入温度为37℃、CO2浓度为5%的培养箱中继续培养。
2.3.2 卵母细胞免疫荧光染色 1)相关试剂的配制。PBS缓冲液:调pH值至7.4,进行高压灭菌,之后保存于4℃;固定液:用4%多聚甲醛配制4%PBS溶液;透膜液(MPs):用TritonX-100配制0.5%PBS溶液;洗脱液(WBF):用Tween 20配制0.1%PBS溶液;封闭液(BBF):用牛血清白蛋白(BSA)配制10%WBF溶液。2)卵母细胞免疫荧光染色方案。收集所需的卵母细胞置于固定液中固定30 min,然后在MPs溶液中透膜30 min,再在硫化新蓝(BBF)溶液中封闭1 h。将封闭后的卵母细胞放入按1∶100BBF溶液稀释的兔源抗PRC1抗体中,4℃孵育12~16 h。之后将卵母细胞转移至WBF溶液中漂洗3遍,每遍5 min。再将卵母细胞转移至按1∶500 WBF溶液稀释后的CyTM5偶联羊抗兔二抗中孵育1 h。之后将卵母细胞转移至WBF溶液中漂洗3遍,每遍5 min。随后将卵母细胞放入按1∶200 WBF溶液稀释后的FITC偶联的小鼠源抗α-Tubulin单克隆抗体中孵育1 h。之后将卵母细胞转移至WBF溶液中漂洗3遍,每遍5 min。结束上述步骤之后,取适量DAPI滴在载玻片中央,将卵母细胞转移到DAPI中,盖上盖玻片,避光保存于-20℃。再用高灵敏度激光共聚焦显微镜观察并拍照。
2.3.3 实时定量聚合酶链式反应 qRT-PCR检测在体外成熟的4个关键时期卵母细胞中PRC1基因mRNA的相对表达水平,每次4个样本,分别为体外培育0,8,10,12 h的卵母细胞,每个样本收集到的卵母细胞数目为60~70个。首先用微量RNA提取试剂盒提取各组样品中的总RNA,之后用逆转录试剂盒进行逆转录反应,得到cDNA,稀释到合适浓度之后用实时定量PCR仪进行qRTPCR反应,以β-actin作为内参基因,最后采用2-ΔΔCt法来对目的基因mRNA相对表达量进行计算,试验至少重复3次。试验数据采用GraphPad Prism 5软件进行处理,并进行单因素方差分析。
3 结果与分析
3.1 PRC1在卵母细胞发育不同时期定位的结果
为了探究PRC1在小鼠卵母细胞减数分裂中的亚细胞定位情况,收集4个关键时期的卵母细胞进行免疫荧光试验,结果见图2。在GV期PRC1定位于生发泡上,在MⅠ、TI和MⅡ期PRC1都定位在整个纺锤体上。
3.2 PRC1基因在不同时期mRNA表达量分析结果
为了进一步研究PRC1基因在小鼠卵母细胞减数分裂过程中表达量的变化,收集4个关键时期的卵母细胞进行qRT-PCR试验,结果见图3。在体外培养8小时时卵母细胞发育到MⅠ期,PRC1基因mRNA表达量与GV期相比稍有上升,但差异不显著(P>0.05);在体外培养10小时时卵母细胞发育到TI期时,PRC1基因mRNA表达量达到最高,与GV期相比差异显著(P<0.05);在体外培养12小时时卵母细胞发育到MⅡ期,PRC1基因mRNA的表达量稍有下降,与GV期相比差异显著(P<0.05)。
4 讨论
哺乳动物的卵母细胞早在胚胎时期就已形成,大多数动物出生前或出生后很短一段时间卵母细胞发育阻滞于第一次减数分裂前期的双线期,此时被称为初级卵母细胞。这一静止期持续时间较长,一直到雌性动物性成熟。雌性动物进入发情期后,在促性腺激素的诱导作用下减数分裂恢复,生发泡破裂,发育至MⅡ期时排出PBI,此时被称为次级卵母细胞。次级卵母细胞发育到MⅡ期时发生了第二次阻滞,这一静止期持续时间较短,一直到精子的到来减数分裂才能进一步进行。与体细胞有丝分裂的不同之处在于,卵母细胞的减数分裂是不均等分裂,产生一个体积较大的卵母细胞和两个体积较小的极体。减数分裂的每个步骤(如生发泡破裂、纺锤体组装、染色体排列、纺锤体迁移和极性的形成等)都涉及到诸多基因的精准调控,只有各步骤精确配合才能保证减数分裂成功。因此,探究每个基因对于该过程可能的影响显得十分重要。
可以通过观察蛋白在细胞中不同位置的表达情况大致推断该蛋白可能在哪些方面发挥着生物学功能。已有研究表明,PRC1是微管结合蛋白,其对纺锤体的形成是十分重要的[7]。本试验证明了在小鼠卵母细胞减数分裂过程中PRC1在GV、MⅠ、TI和MⅡ期都稳定表达,且与纺锤体微管处于共定位的状态。该结果说明PRC1可能会与其他的纺锤体微管蛋白相互作用,从而调节卵母细胞减数分裂的过程。可以通过观察基因在不同时期的mRNA表达量大致推断该基因调控着怎样的生理过程。之前的研究发现,在癌细胞中下调PRC1表达量会造成癌细胞发育阻滞,最终导致细胞凋亡[8],说明PRC1基因表达量的不同可能会造成不同生物学功能。本试验结果表明,以GV期卵母细胞中PRC1基因mRNA表达量为对照,随着小鼠卵母细胞体外发育的进程,PRC1基因mRNA表达量逐渐上升,在小鼠卵母细胞体外培养到达TI期时基因表达量最高,只是发育到了MⅡ期之后出现了轻微下调。说明PRC1在TI期的高表达可能有促进第一极体排出的作用。
综上所述,PRC1在小鼠卵母细胞减数分裂过程中与纺锤体处于共定位的状态,并且在TI期PRC1基因mRNA水平最高,表明PRC1可能在第一极体排出的过程中发挥着重要作用。结合前期其他人的研究报道,推测PRC1可能与Rho家族成员之一GTP激动蛋白酶(MgcRac-GAP)相互作用调节减数分裂的过程。与此同时,在卵母细胞中PRC1与KIF4、KIF14、胞质连接蛋白1等纺锤体微管形成相关蛋白对于减数分裂的调控机制也值得深入探讨。此外,通过查阅相关文献发现,PRC1在家畜卵母细胞减数分裂过程中的调控机制尚不明确。因此关注PRC1在家畜生殖细胞中的作用机制具有更为重要的意义。
