养殖池塘中3种常见水草的叶绿素荧光特征比较
2020-07-06陶菊红韩晓磊袁静雯曹亚男张涛
陶菊红 韩晓磊 袁静雯 曹亚男 张涛


摘要:以养殖池塘中常见的3种沉水水草为材料,通过调制荧光技术研究了不同物种水草的叶绿素荧光特征。结果表明:在给定的光照度下[130 μmol/(m2·s)],伊乐藻和金鱼藻实际量子效率(ΔF/F′m)、光化学猝灭系数(qP)均显著高于苦草;在快速光曲线参数中,伊乐藻和金鱼藻相对电子传递速率(rETR)、快速光曲线初始斜率(α)、半饱和高光强(Ik)和最大相对电子传递速率(Pm)也显著高于苦草;诱导曲线参数非光化学淬灭系数(NPQ)的结果与之有所不同,金鱼藻和苦草相似且均高于伊乐藻样品。结果表明,不同水草物种具有各自的叶绿素荧光特征,其中伊乐藻和金鱼藻对光能的利用效率相对较高,伊乐藻能够适应相对高的光照条件,而苦草则更适合在较低光照条件下培植。
关键词:水草;养殖池塘;叶绿素荧光;光照
中图分类号:S917.3 文献标志码: A 文章编号:1002-1302(2020)10-0198-04
收稿日期:2020-01-14
基金项目:苏州市科技计划(编号:SNG2017073)。
作者简介:陶菊红(1981—),女,江苏苏州人,博士,助理研究员,主要从事植物学研究。E-mail:taojuhong@126.com。
通信作者:张 涛,博士,副研究员,主要从事植物学研究。E-mail:zhangtofy@163.com。
水草的人工栽培在水产养殖中起到重要作用,是池塘生态养殖的关键措施[1-3]。人工栽培水草在池塘养殖中具有多重功能,其一是可以作为养殖动物的天然饵料,常见的有伊乐藻(Elodea nattalii)、苦草(Vallisneria spiralis)、金鱼藻(Ceratophyllum demersum)、轮叶黑藻(Hydrilla verticillata)等[3-4];其二是水草的栽培能够改善和提高养殖水体环境,通过吸收水体中的有害物质,有效阻止水体富营养化,还可通过光合作用生成氧气以供养殖鱼虾蟹等的生长[5-7];其三是能够为养殖动物提供栖息场所,使鱼、虾、蟹等能够顺利完成生长发育的关键阶段[2,4,8]。
随着社会的发展,水产养殖过程中越来越注重生态元素,循环养殖技术也得到大力发展,其中养殖池塘中水草的应用是净化水质的有效手段,得到从业者及水产科研人员的关注。江苏省目前主要栽培的水草物种比较多,常见的有伊乐藻、苦草、金鱼藻、水花生等[2],这些水草具有不同的生物学特征。不同的水草在养殖池塘中所起的作用也有差异,生产实践表明,作为沉水性水生维管束植物,伊乐藻、金鱼藻等对温度的适应能力较强,往往是池塘养殖的优势水草物种;苦草可供水生动物攝食,能够起到天然饵料和水质调节的双重作用[2,9-10]。因此,如何根据水草的物种特性进行科学的组合栽培,充分发挥水草在池塘养殖领域的生态作用已成为水产生态养殖中的重要课题。现有的研究主要集中在水草的栽培数量、物种等方面,保持适宜的水草生物量能够维持良好的水质[10],在降低养殖虾蟹等发病率的同时,还能够提高其品质和产量[8]。徐增洪等研究表明,不同物种水草合理搭配对池塘养殖生态的调节作用更佳,而且认为水草栽培面积应进行适当的人为控制[4]。
尽管人工移植水草在池塘生态养殖方面的重要性得到越来越多的重视,但是生产上仍存在一定的问题,例如多数养殖池塘以水草自然生长为主,而且选用水草物种单一或者组合效果不佳等等,其中一个重要的原因是对水草生物学特征的研究较少。养殖池塘中水草的生长状态与其光合生理活性密切相关,因此研究不同水草物种光合生理过程,将有助于优化栽培物种的组合以及控制其生物量。
1 材料与方法
1.1 供试材料
于2018年10月13日江苏省苏州市相城区虾蟹养殖池塘中采集3种人工主要栽培水草:伊乐藻、金鱼藻和苦草,将样品采集后置于水桶中随即送至实验室处理。首先用自来水清洗,除去表面杂质,随后选取健康水草置于干净水体中,于自然光照,水温为 20 ℃条件下,待进行叶绿素荧光测定。
1.2 叶绿素荧光特征测定
采用叶绿素荧光仪(Water-PAM,德国)测定样品。所设参数如下:
实际量子效率(ΔF/Fm′):将光纤探头垂直对准水草叶片表面进行测定。参照培养时环境光照强度,首先提供0.3 μmol/(m2·s)调制红光和 130 μmol/(m2·s) 活化光,稳定后即测得样品实时荧光值Ft[11];随后提供0.8 s、4 000 μmol/(m2·s)左右的饱和脉冲光,测得样品最大荧光值Fm′。将所得荧光值代入以下方程计算光系统Ⅱ(PSⅡ)的实际量子效率:
ΔF/Fm′=(Fm′-Ft)/Fm′。
快速光曲线(RLC):设定仪器连续为样品提供8个光照度:90、140、210、310、480、710、1 000、1 400 μmol/(m2·s)。每个光照度持续照射20 s,以保证Ft能够达到稳态,随后施加0.8 s的 4 000 μmol/(m2·s) 饱和脉冲光,测得此时样品的最大荧光值Fm′。
相对电子传递速率(rETR)计算公式如下:
rETR=ΔF/Fm′×PFD×0.5×0.84。
式中:系数0.84表示环境光量子的84%被样品吸收,PFD是光子通量密度。
以PFD和rETR的值分别作为x、y轴作图,即得到快速光曲线(RLC)。参照Platt等的方法[12]对RLC进行拟合,计算出初始效率(α),最大相对电子传递速率(Pm)和半饱和光强(Ik)等参数,计算公式如下:
P=Pm(1-e-αPAR/Pm)·e-βPAR/Pm;
Ik=Pm/α。
式中,PAR为光照度,β为光抑制参数。
诱导曲线:首先将样品进行暗适应30 min,目的是使得PSⅡ处于完全开放状态。之后打开测量光[光照度约0.3 μmol/(m2·s)],测得最小荧光值(Fo),然后提供0.8 s的4 000 μmol/(m2·s)饱和脉冲光,此时得到最大荧光值(Fm),随后关闭光源,40 s后打开活化光[光照度138 μmol/(m2·s)],之后每隔20 s照射1次与之前相同强度的饱和脉冲光,直至最大荧光值(Fm′)和最小荧光值(Fo′)均达到稳定状态[13]。采集得到的荧光值经以下公式计算光化学猝灭(qP)和非光化学猝灭系数(NPQ):
qP=(Fm′-F)/(Fm′-Fo′);
NPQ=(Fm-Fm′)/Fm′。
1.3 统计分析
所有试验中,均设3~5个重复,试验数据表示为平均值±标准差(n≥3),对数据结果进行t检验,以0.05为显著差异水平。
2 结果与分析
2.1 不同样品实际量子效率
试验给定的130 μmol/(m2·s)活化光下,不同水草的实际量子效率(ΔF/Fm′)存在差异(图1),其中伊乐藻和金鱼藻样品的ΔF/Fm′相差不大,均在0.45左右,苦草样品ΔF/Fm′最低,约为0.08,显著低于伊乐藻和金鱼藻样品(P<0.05)。
2.2 不同样品快速光曲线结果
由图2可知,随着光照度的上升,所有样品的相对电子传递速率均表现为低光照度下快速上升,待达到一定光照度时保持较为稳定的变化特点,这符合光合植物的rETR特征。
不同水草RLC中rETR数值存在差异,相同光照条件下,金鱼藻rETR最高,伊乐藻样品次之,苦草样品rETR显著低于另外2种水草样品(P<0.05);而且,金鱼藻和伊乐藻样品rETR在光照度为 500~700 μmol/(m2·s) 时达到最大,而苦草rETR在 100 μmol/(m2·s) 左右达到最大值。
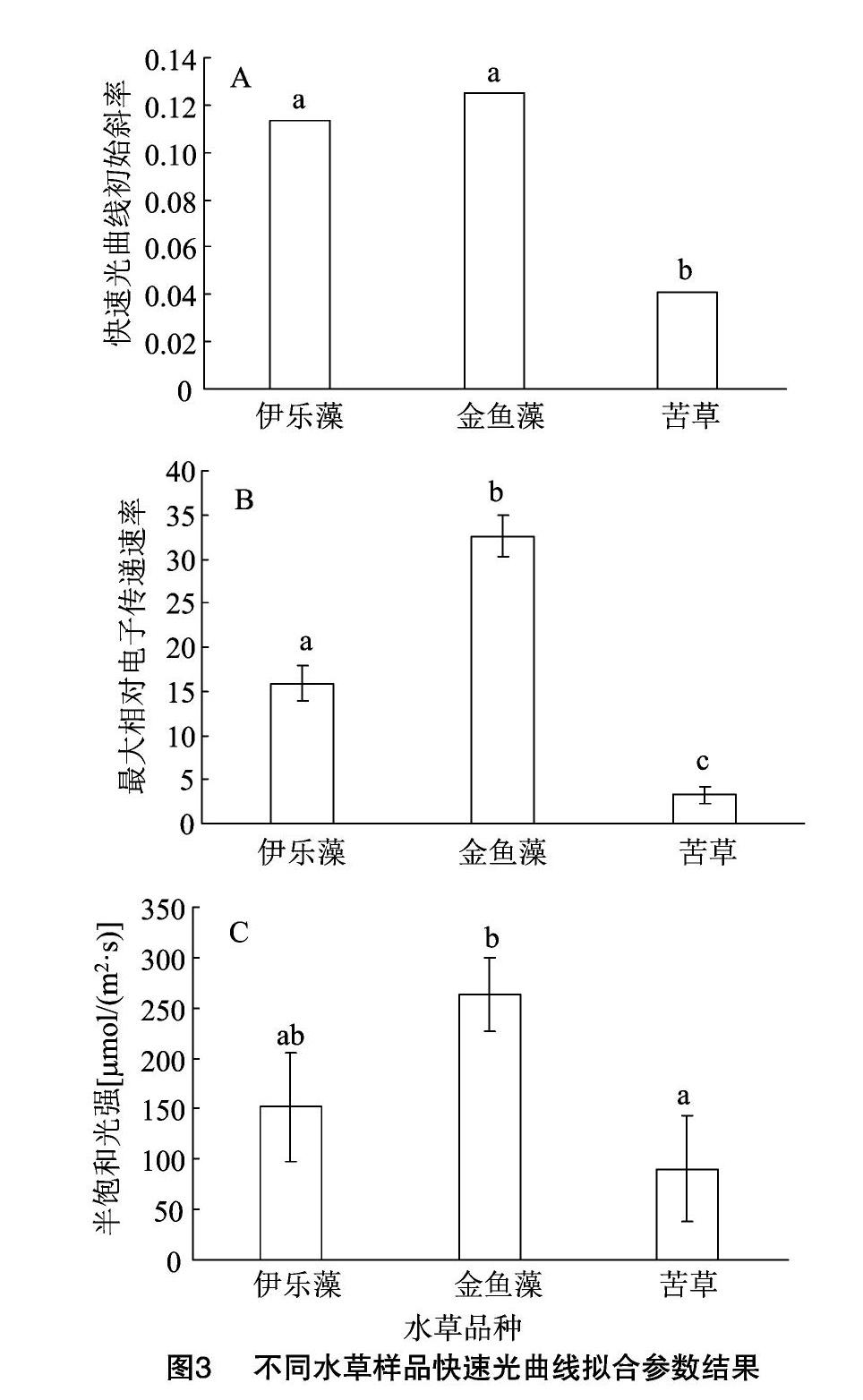
2.3 快速光曲线参数结果
伊乐藻和金鱼藻样品RLC的初始斜率差异不明显(图3-A),但均显著高于苦草。最大相对电子传递速率(Pm)的计算结果显示(图3-B),金鱼藻样品Pm值最高,约为32,伊乐藻样品Pm值15左右,苦草样品Pm值最低,仅为5左右。半饱和光强(Ik)的计算结果(图3-C)显示,金鱼藻的Ik值最高,伊乐藻次之,而苦草最低,其中金鱼藻的Ik显著高于苦草。
2.4 诱导曲线参数
图4结果表明,在给定光照度为 138 μmol/(m2·s) 的条件下,伊乐藻和金鱼藻的光化学猝灭系数(qP)在0.7左右,均显著高于苦草;而非光化学猝灭系数(NPQ)的结果则与之不同,金鱼藻和苦草的NPQ较高,而伊乐藻最低。
3 讨论
在池塘养殖过程中,已报道了人工栽培的多种水草物种,其中常见的沉水水草物种主要有伊乐藻、轮叶黑藻、金鱼藻、苦草等[2,9]。这些水草物种因其原产地等原因表现出不同的环境适应特征。伊乐藻原产美洲,对环境有着较强的适应能力,在春季、晚秋及冬季均能较好地生长繁殖,而夏季随着水温的升高,伊乐藻的生长受到抑制,严重高温条件甚至会导致其死亡;金鱼藻在水产养殖的后期即初秋季节能够成为占生长优势的物种;苦草主要分布于温带、热带地区,喜相对较高的温度,同时还是河蟹喜欢的天然食物[2,10]。本试验结果表明,晚秋时间采集的3种水草物种叶绿素荧光特征存在差异,这与其原产地环境特征密切相关,试验期水温20 ℃左右,此时伊乐藻在较高的光照度下能够保持较高的实际量子效率和光化学淬灭系数,同时也具备较高的半饱和光强,说明在这一季节,伊乐藻对光照条件有着更好的适应能力;本试验中金鱼藻能够在给定的光照度下保持较高的实际量子效率,但非光化学猝灭系数较高,意味着此时金鱼藻需要通过对吸收的光能进行调节分配,来提高热耗散以适应光照度的升高;在本试验条件下,苦草的光能利用效率处于比较低的水平,表明与其他2个水草物种比较,苦草更适应较低的光强环境,光照度过高会导致苦草对光能的利用效率降低,同时也明显提高了热耗散所占的比例。
养殖池塘除随季节而产生的水温变化外,水体的光照条件也是关键的环境因素。实际生产表明,养殖池塘水体的透光度受多种因素影响,一般是通过施加足量饵料来调节透光度,此时养殖虾蟹觅食活动会搅浑池水,降低透光度;此外施加微生物制剂,减少腐生细菌的大量繁殖等也可调节水体的透光度[2],然而这些方法的使用均会影响水草生长环境的光照条件。并且水草如果过量生长或栽植除了影响鱼虾蟹的生长外[3-4,8],也会降低水体的透光度,因此需要人为进行打捞以保证较好的生长环境和光照条件,以发挥更好的生态积极效用。对3种沉水水草的叶绿素荧光特征研究可推知,伊乐藻能够适應较强的光强,可以栽培在透光度较好的区域,金鱼藻能够通过调节对吸收光能的分配以适应环境光强的变化,可以主要栽培在透光度中等的区域,而苦草更适应低光照条件,透光度较低的区域是其生长的适宜区域。
在养殖池塘中水草的人工栽培是影响鱼虾蟹生长的重要环境因素,通过对其光合生理的研究将有助于更清楚地了解不同水草物种的适宜栽培条件,包括栽培季节、栽培区域等,进而可以通过物种的良好搭配,形成适宜的生长环境,促进养殖池塘中鱼、虾、蟹的优质生长。
参考文献:
[1]宋长太. 水草在河蟹养殖中的作用及栽培[J]. 北京水产,2000(6):18-19.
[2]谢国兴,陈正锦,鲍胜华,等. 河蟹生态健康养殖池塘中水草的栽培[J]. 水产养殖,2013(12):45-47.
[3]李 伟. 鱼虾蟹类健康养殖中水草的作用分析[J]. 南方农业,2017,11(32):78-79.
[4]徐增洪,刘国锋,水 燕,等. 人工栽培水草对池塘养殖生态环境和虾蟹生长的影响[J]. 江苏农业科学,2016,44(6):328-331.
[5]李文朝. 富营养水体中常绿水生植被组建及净化效果研究[J]. 中国环境科学,1997,17(1):55-59.
[6]Petrucio M M,Esteves F A. Uptake rates of nitrogen and phosphorus in the water by Eichhornia crassipes and Salvinia auriculata[J]. Revista Brasileira de Biologia,2000,60(2):229-236.
[7]Qiu D R,Wu Z B,Liu B Y,et al. The restoration of aquatic macrophytes for improving water quality in a hypertrophic shallow lake in Hubei Province,China[J]. Ecological Engineering,2001,18(2):147-156.
[8]李定国,王煜恒,王会聪,等. 水草密度对河蟹池塘水质和养殖效益的影响[J]. 水产养殖,2015(12):11-15.
[9]岳帮亚. 水草在河蟹生态健康养殖中的应用[J]. 现代农业科技,2010(17):334-335.
[10]刘英钦,管雪婷. 浅谈养蟹池塘中的水草种植[J]. 畜牧獸医科技信息,2018(10):16-17.
[11]张 涛,沈宗根,姚春燕,等. 基于叶绿素荧光技术的紫菜光适应特征研究[J]. 海洋学报,2011,33(3):140-147.
[12]Platt T,Gallegos C L,Harrison W G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton[J]. Journal of Marine Research,1980,38(4):687-701.
[13]李家富,张 涛,陆勤勤,等. 不同温度条件下条斑紫菜叶绿素的荧光特征[J]. 江苏农业科学,2014,42(1):200-203.
