屋顶绿化基质及植物耐旱性研究
2020-07-06冯晓燕殷金岩汪田野王东栋樊腾新梁令帅裴福云
冯晓燕 殷金岩 汪田野 王东栋 樊腾新 梁令帅 裴福云
摘要:以小蚌兰、翠芦莉及颗粒基质为材料,开展颗粒基质保水性能的蒸发试验及盆栽植物控水干旱试验,以获得相关性能指标[不同干旱处理下(0、7、14、21 d)基质含水量、植物叶片相对含水量、叶绿素含量(SPAD值)、凈光合速率、蒸腾速率、相对电导率、丙二醛(MDA)含量及根系生物量等指标,并采用隶属函数法对不同处理各指标进行综合评价分析],探究自主研发的轻型颗粒基质抗旱保墒性能及相关植物的耐旱性。结果显示,颗粒基质具有良好的吸水、保水性能,在干旱条件下能为植物提供较为适宜的基质生长条件;小蚌兰在同等干旱条件下比翠芦莉更具有耐旱性。本研究将为颗粒基质抗旱保墒性能的评价及轻质、节水型屋顶绿化景观的建立提供科学依据。
关键词:屋顶绿化;干旱胁迫;颗粒基质;小蚌兰;翠芦莉;基质含水量;叶片生理指标;综合评价;隶属函数值
中图分类号: S152;S688 文献标志码: A 文章编号:1002-1302(2020)10-0162-06
收稿日期:2019-05-12
基金项目:广东省深圳市节能环保产业发展专项(编号:深发改[2016]1023号)。
作者简介:冯晓燕(1989—),女,河南信阳人,硕士,中级工程师,主要从事植物废弃物资源化利用、立体绿化等研究。E-mail:elain2016@163.com。
通信作者:裴福云,博士,工程师,主要从事纳米材料在土壤修复、光催化领域方面的应用及新型基质材料研究。E-mail:peify@126.com。
随着城市化进程加快,屋顶绿化作为一种新型的绿化方式,不仅可以弥补平地绿化的不足,缓解建筑面积和绿地面积之间的矛盾,还可以改善人居环境,彰显出不可低估的生态、社会及经济效益[1-2]。但屋顶环境较为特殊,屋顶绿化不仅要考虑承重,还须考虑强光照、大风、极端温度等恶劣的环境条件,而且基质不与地表相连,易缺水干旱[3]。而干旱一直是制约植物生长发育的主要逆境因子,在园林植物选材日益丰富的情况下,抗旱品种的供应需求也日趋旺盛[4]。鉴于栽培基质占屋顶绿化系统质量比重较大,对植物的生长也影响最大,因此对基质的要求更高[5],可见选择吸水、保水性能好、轻质的基质材料至关重要。本研究通过试验探究自主研发的轻型颗粒基质的吸水、保水性能及耐旱性,并研究2种不同植物对基质及干旱胁迫的适应性,旨在为建立轻质、节水型屋顶绿化景观、实现城市可持续发展提供科学依据。
1 材料与方法
1.1 材料
1.1.1 试验材料与试剂 供试材料包括颗粒基质、普通营养土、植物(翠芦莉、小蚌兰)、自封袋、信封、滤纸、花盆、直尺等。试剂包括三氯乙酸(TCA)、2-硫代巴比妥酸(TBA)等,均购自东莞市常平正科化玻仪器店。
1.1.2 仪器与设备 培养皿、镊子、研钵、分析天平、筛子、离心管,均购自东莞市常平正科化玻仪器店;移液枪、便携气象计、紫外分光光度计、电导仪、手持便携式叶绿素仪、Li-6400全自动便携式光合测量仪等。
1.2 试验方法
1.2.1 室内常温常压蒸发试验 在关闭门窗(以保持室内无风状态且其温湿度变化较小)的环境条件下进行试验。颗粒基质为试验组,普通基质土为对照,每组3个平行试验。试验中使用开口培养皿为蒸发器皿,以精度为0.01 g的电子天平和电子温湿度表为称量仪器,具体试验方法如下:称量一定体积颗粒基质及普通基质土为试验材料,使试样达到饱和,将饱和后的试样均匀摊铺在培养皿内,记录时间、温度和相对湿度以及试样和蒸发器皿的总质量。每 3 h 称量蒸发器皿质量、温度和相对湿度,直至蒸发过程结束[6]。
1.2.2 盆栽控水干旱试验及指标测定 试验于2018年6—7月进行,在广东省东莞市铁汉生态农业观光园的抗逆植物大棚(114.10°E、22.99°N)内开展盆栽控水干旱试验,花盆规格为15 cm×15 cm×18 cm,试验组(T)所用基质为自制颗粒基质,对照组(CK)为普通营养土,分别作控水(T干、CK干)和正常浇水养护(T水、CK水)2组处理。选取株龄长势大小一致的翠芦莉、小蚌兰植株定植于试验组及对照组花盆中,定植密度为1株/盆。控水组在6月下旬进行干旱胁迫,在控水前1 d浇透水,使每盆的基质相对含水量基本饱和,然后开始控水,连续控水21 d;浇水组每周浇水1次。分别于控水0、7、14、21 d测定土壤指标及植物光合和生理生化指标。
基质含水量、叶片相对含水量及根系生物量采用烘干法[7]测定;叶绿素含量(以SPAD值计)采用手持便携式叶绿素仪(SPAD-502,Japan)测定;相对电导率采用电导仪[7]测定;光合指标采用Li-6400全自动便携式光合测量仪测定;丙二醛(MDA)含量采用硫代巴比妥酸法[8]测定。
2 结果与分析
2.1 室内常温常压蒸发试验结果分析
基质水分蒸发是基质内部水分经过基质表面以水蒸气的状态扩散到大气的过程。影响基质水分蒸发的因素有很多,归纳起来可分为2类:内因和外因。内因是指与基质自身相关的各类化学物理性质因素,主要影响水分在基质内的运移特征。外因是指基质蒸发时的外部环境因素,主要影响液态水汽化所需要的能量供应强度[9]。本试验各个处理组处于同一环境条件下,变动幅度不大,所以影响基质蒸发的主要因素为内因。
如图1所示,随着蒸发时间的延长,前期基质水分蒸发速率较大,然后逐渐降低,这主要是因为前期基质蒸发的水分主要是毛管水,较容易蒸发,然后是薄膜水和吸着水,在同等条件下几种水分蒸发越来越难。由表1可知,同等体积的基质,颗粒基质初始含水量(以质量分数计,下同)是普通基质土的4.8倍,蒸发9 h后,颗粒基质含水量仍高达98.70%,而普通基质土仅有12.07%;室内蒸发50 h后,普通基质土含水量仅剩0.20%,基本蒸发完全,而颗粒基质为589%。由此可以看出,在同等蒸发时间及蒸发条件下,颗粒基质具有良好的吸水、保水性能。
2.2 盆栽控水干旱试验各指标分析
2.2.1 干旱胁迫对基质含水量的影响 一般随着
干旱胁迫加剧,基质含水量会呈不同程度的下降趋势。由表2可知,随着胁迫时间的延长,控水干旱胁迫下试验组及对照组基质含水量均呈持续下降趋势,前期下降速率快,后期较为缓慢。其中,颗粒基质饱和含水量较高,能达基质自身干质量的近15倍。颗粒基质(T)含水量在持续干旱(控水)过程中一直高于CK普通营养土,干旱21 d后,小蚌兰的T干旱组比CK干旱组基质含水量高5.31百分点,翠芦莉T干旱组比CK干旱组基质含水量高2.36百分点,T浇水组基质含水量也远远高于CK浇水组,说明颗粒基质具有良好的吸水、保水性能,且颗粒基质材质轻,更符合屋顶绿化承重要求。通过颗粒基质T干旱组数据可以看出,小蚌兰基质含水量持续高于翠芦莉;CK干旱组的小蚌兰的基质含水量胁迫0~14 d均高于翠芦莉组,21 d时接近持平,由此推测小蚌兰抗旱性优于翠芦莉。
2.2.2 干旱胁迫对叶片相对含水量的影响 一般情况下,植物在受到干旱胁迫时,会导致叶片水分亏缺,相对含水量会呈下降趋势,通常其下降的速率与抗旱能力呈负相关关系。植物在遭受同样的干旱时,抗旱性越强的植物叶片相对含水量下降的幅度越小;抗旱性越差的植物叶片相对含水量下降幅度越大[10]。由表3可以看出,浇水组基本上处于动态平衡过程。干旱组在持续水分胁迫下, 小蚌兰与翠芦莉叶片相对含水量整体呈持续下降趋势,颗粒基质干旱组胁迫7 d后叶片相对含水量基本与饱和含水量持平,总体下降趋势较为平缓,但CK干旱组下降速率快,干旱胁迫21 d,小蚌兰及翠芦莉CK干旱组叶片相对含水量分别比T干旱组低23.62、31.08百分点,可推测颗粒基质在干旱条件下更适宜植物生长。干旱胁迫21 d后,T干旱组的小蚌兰叶片相对含水量下降1.86百分点,翠芦莉下降3960百分点;CK干旱组的小蚌兰下降25.50百分点,翠芦莉下降71.07百分点,说明小蚌兰在干旱条件下自身反馈调节能力强,能够更好地适应于干旱胁迫。
2.2.3 干旱胁迫对叶绿素含量的影响 由于植物在遭受干旱胁迫时其叶片叶绿素含量关系着植物的光合同化过程,所以叶绿素含量是衡量植物抗旱性的重要生理指标之一[11]。一般在遭受到干旱胁迫时植物的叶绿素含量都呈下降趋势,所以通过测量各处理植物在干旱胁迫时叶绿素含量可以反映出不同种类植物对干旱胁迫的响应机制。由表4可以看出,随着干旱胁迫加剧,小蚌兰的叶绿素含量呈持续下降趋势,T干旱组胁迫期内降低 5.4,且CK干旱組胁迫后期,降幅较大,胁迫期内降低132,可推测干旱胁迫下,颗粒基质因保水性更好,能为植物提供适量水分,缓解叶绿素分解速度;而翠芦莉T干旱组则呈“升高—降低—升高”趋势,分析原因可能是干旱胁迫初期,叶片内部保护机制使得叶绿素含量减少,并且叶片中含水量降低,导致叶绿素含量反而增加,中期叶绿素分解加剧导致含量降低,后期叶片发生萎蔫,叶绿素浓缩导致含量增加。翠芦莉CK干旱组呈持续上升的趋势,由此推测,随着水分胁迫的持续,叶片含水量降低,而叶片叶绿素含量因叶绿素浓缩而持续上升。
2.2.4 干旱胁迫对植物相对电导率的影响 叶片细胞膜相对透性(相对电导率)是生物膜系统能否稳定的重要指标,膜系统稳定是细胞进行正常生理功能的基础。植物在遭受到干旱胁迫后,叶片细胞膜受到损伤,质膜透性会增加,细胞内含物外渗,电导率会增大[12]。如表5所示,随着水分胁迫加剧,植物叶片的相对电导率均呈不同程度的动态变化,其中小蚌兰T干旱组呈持续上升趋势,胁迫后21 d
相对电导率增加38.35百分点,CK干旱组呈先升后降趋势,可能是植物自身对干旱胁迫的调节机制,对干旱有一定的适应性,叶片相对电导率也随之小幅度下降,最终(胁迫后21 d)增加41.64百分点,比T干旱组多增加3.29百分点;翠芦莉的T干旱组与CK干旱组相对电导率均呈“升高—降低—升高”趋势,在土壤水分胁迫前期相对电导率升高,一段时间后,植株对胁迫形成了一定的抗性,相对电导率降低,但随着胁迫时间延长和程度加剧,植物自身调节幅度达到极限,导致相对电导率再次升高。
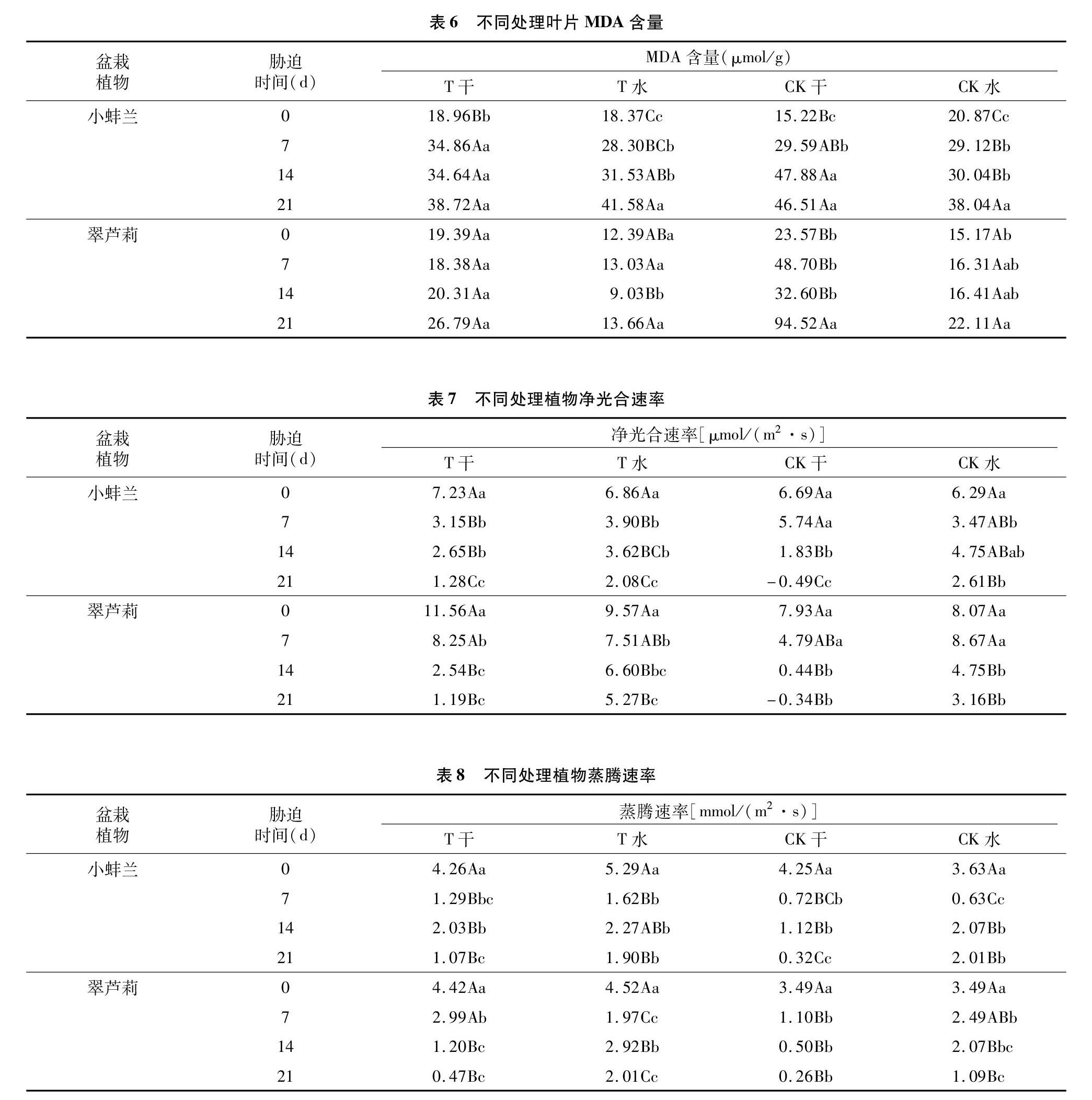
2.2.5 干旱胁迫对MDA含量的影响 植物在遭受到干旱胁迫时会导致大量的自由基产生,致使MDA生成。一般来说,在干旱胁迫下,叶片MDA含量越高,其抗旱性越差[13]。由表6可以看出,在水分胁迫期内,不同处理的MDA含量均整体呈不同程度的增加,其中小蚌兰T干旱组在胁迫后21 d时增加19.76 μmol/g,CK干旱组增加31.29 μmol/g;翠芦莉T干旱组增加7.40 μmol/g,CK干旱组增加70.95 μmol/g,CK干旱组的增幅比T干旱组多6355 μmol/g。可见,颗粒基质因保水性较好,植物在同等水分胁迫条件下,产生的MDA也较少。
2.2.6 干旱胁迫对净光合速率的影响 植物在遭受到干旱胁迫时,净光合速率会下降,下降的幅度与植物的抗旱性呈正相关[14]。由表7可以看出,干旱处理组小蚌兰和翠芦莉净光合速率均呈持续下降趋势;CK干旱组胁迫21 d后,小蚌兰、翠芦莉净光合速率均出现负值,极度缺水会抑制光合作用,T干旱组净光合速率仍为正值,说明颗粒基质更能适应干旱环境,更能为植物在干旱环境中提供生存条件。浇水组净光合速率也会一定程度降低,但植物能保持正常的生长。
2.2.7 干旱胁迫对蒸腾速率的影响 植物的蒸腾作用在植物水分代谢中起着很重要的调节支配作用,随着干旱胁迫加剧,通过气孔关闭来减少水分散失。由表8可以看出,随着干旱胁迫的持续,小蚌兰不同处理蒸腾速率呈“降低—升高—降低”趋势,原因可能是干旱胁迫7 d时,刚雨过天晴,空气湿度较大,蒸腾速率低;而翠芦莉干旱组持续下降,浇水组则呈“降低—升高— 降低”趋势。干旱胁迫21 d后,小蚌兰的T干旱组蒸腾速率降低3.19 mmol/(m2·s),CK干旱组蒸腾速率降低3.93 mmol/(m2·s),说明颗粒基质优于普通营养土; 翠芦莉组分别降低 3.95、3.23 mmol/(m2·s),说明普通营养土则优于颗粒基质。
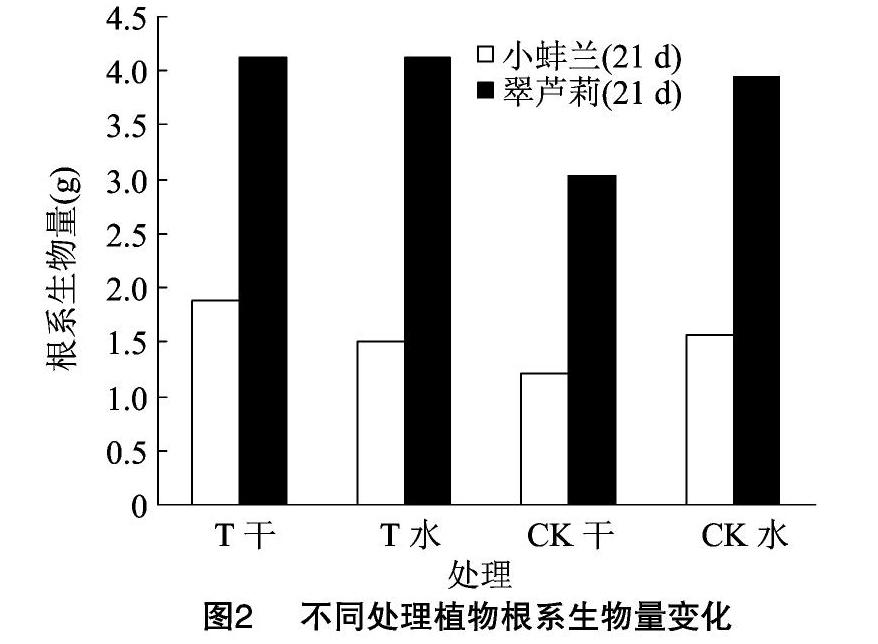
2.2.8 干旱胁迫对根系生物量的影响 一般植物在缺少水分的环境下,会形成较大的根冠比,以充分吸收、利用水分,维系正常生理活动。如图2所示,干旱胁迫21 d后,干旱组与对照组根系生物量差异不显著,但小蚌兰和翠芦莉T干旱组根系生物量值大于CK干旱组,结合表2可以看出,T干旱组基质因含水量高于CK干旱组,有利于植物根系生长,另外颗粒基质团粒结构有利于根系穿插生长。通常建筑屋顶的承重有限,用于屋顶绿化的土壤基质往往特别浅薄(2~15 cm),因而在屋顶绿地环境中植物的根系生长所必需的资源与其发展所必需的空间十分有限,因而大体量的根系系统也不适合在屋顶基质中生长[3]。故翠芦莉这种根系发达、根系体量大的植物也不适宜于屋顶种植,以免争夺资源而对屋顶产生破坏性影响。
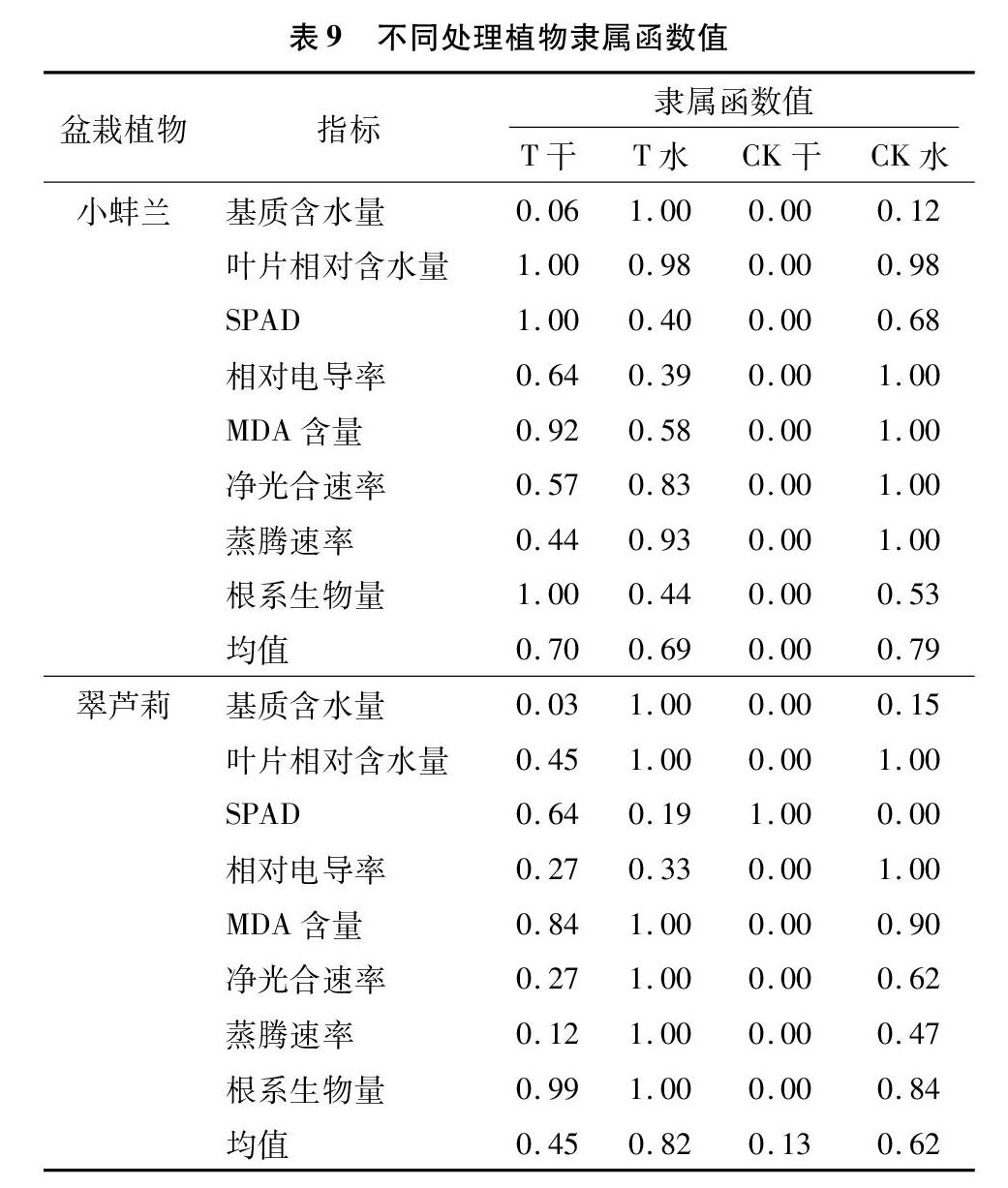
2.3 盆栽控水干旱试验各指标综合分析评价
由于抗旱性受多因素相互作用、相互影响,单一的抗旱性指标往往难以全面反映植物对干旱适应
的综合能力,所以采用隶属函数综合评价法能够消除单个指标带来的片面性,以对比各物种抗旱差异性[15],故采用隶属函数法对不同处理干旱胁迫21 d的生长指标进行综合分析评价。由表9可以看出,小蚌兰的不同处理中CK浇水组值最高,各处理依次表现为CK水>T干>T水>CK干,干旱组的颗粒基质优于CK普通土,说明对于小蚌兰而言,使用颗粒基质在低浇水量条件下就可以接近普通土在高浇水量条件下的生长水平。翠芦莉不同处理中T浇水组值最高,各处理依次表现为T水>CK水>T干>CK干。在澆水状态下,小蚌兰的CK优于T,干旱状态下的T处理仅次于CK水;对于翠芦莉,浇水组优于干旱组,干旱状态下T优于CK。可见,颗粒基质在干旱条件下的性能优于普通基质土,因具有良好的吸水、保水性能,能为植物提供良好的土壤生长环境,在干旱环境条件下也表现优异。而小蚌兰的T干旱组隶属函数值为0.70,翠芦莉为045,说明小蚌兰耐旱性优于翠芦莉,更能适应屋顶干旱缺水的环境条件。
3 讨论与结论
屋顶绿化由于屋顶承重限制及其特殊的环境条件,所需栽培基质较普通绿化有着不同的要求。轻质保水型屋顶绿化栽培基质能够直观地减少荷载,对屋顶绿化的建设有直接的推动作用[16]。本研究以自主研发的轻型颗粒基质为研究对象,通过颗粒基质的室内蒸发试验及盆栽控水干旱试验可以看出,当干旱胁迫程度及时间呈梯度变化时,基质指标、植物的生理生化指标及光合指标会出现不同趋势及程度的变化,且各指标会相互作用、相互影响,并对耐旱性产生影响。通过基质蒸发试验可知,颗粒基质相对于普通基质土而言,具有良好的吸水、保水性能,在干旱条件下能为植物提供较为良好的土壤条件。
通过小蚌兰和翠芦莉2种不同植物在干旱胁迫和正常浇水养护条件下的各指标变化趋势及幅度可得出,小蚌兰的抗旱性要强于翠芦莉,翠芦莉在干旱胁迫21 d时,已出现净光合速率为负值、植物叶片严重萎蔫现象,且根系较为庞大,可能会对屋顶结构层产生破坏性影响,故不适宜用于基质层较浅、管养频率过低的屋顶绿化工程。
由于华南地区易遭受台风暴雨,干旱试验在大棚内,采用盆栽控水法开展研究,故所测试的植物会受花盆容积的限制,不能广泛扎根,并且与室外屋顶环境不一致,该研究只能做初步对比研究,日后还须开展屋顶试验进一步验证。
参考文献:
[1]汤 聪,刘 念,郭 微,等. 广州地区8种草坪式屋顶绿化植物的抗旱性[J]. 草业科学,2014,31(10):1867-1876.
[2]史晓松,钮科彦. 屋顶花园与垂直绿化[M]. 北京:化学工业出版社,2011.
[3]杨 静. 华南地区四种鸭趾草科轻型屋顶绿化植物的抗旱耐热性研究[D]. 广州:仲恺农业工程学院,2017.
[4]王莺璇. 7种百合科园林地被植物的抗旱性研究[D]. 昆明:云南农业大学,2012.
[5]和晓艳. 屋顶绿化的相关技术研究[D]. 南京:南京林业大学,2013.
[6]张 华,李 茂,曹金露,等. 常用屋顶绿化基质材料的保水性能[J]. 土木建筑与环境工程,2015,37(1):67-72.
[7]郝建军,康宗利,于 洋. 植物生理学实验技术[M]. 北京:化学工业出版社,2006:23-169.
[8]张志良,李小芳. 植物生理学实验指导[M]. 北京:高等教育出版社,2016.
[9]唐朝生,施 斌,顾 凯. 土中水分的蒸发过程试验研究[J]. 工程地质学报,2011,19(6):875-881.
[10]刘成名. 石漠化治理区抗旱性牧草筛选种植技术与示范[D]. 贵阳:贵州师范大学,2015.
[11]孙 龙,彭祚登,王 冲,等. 干旱胁迫对两种沙地灌木能源树种光合特性的影响[J]. 南京林业大学学报(自然科学版),2014,38(2):99-104.
[12]何 玮,蒋 安,王 琳,等. PEG干旱胁迫对红三叶抗性生理生化指标的影响研究[J]. 中国农学通报,2013,29(5):5-10.
[13]石永红,万里强,刘建宁,等. 干旱胁迫对6个坪用多年生黑麦草品种抗旱性的影响[J]. 草地学报,2009,17(1):52-57.
[14]王 强,陈存根,钱红格,等. 水分胁迫对6种苗木光合生理特性的影响[J]. 水土保持通报,2009,29(2):144-149.
[15]张寅媛,刘 英,白 龙. 干旱胁迫对4种景天科植物生理生化指标的影响[J]. 草业科学,2014,31(4):724-731.
[16]王啸宇,郑思俊,张青萍,等. 屋顶绿化适用栽培基质材料保水特性比较研究[J]. 西北林学院学报,2017,32(5):257-262.
