退化喀斯特森林不同植物叶片气孔导度对环境因子的响应
2020-07-04丁晓雅罗绪强冯双任艳莫愁陆汉芝李艳琴秦会斌
丁晓雅 罗绪强 冯双 任艳 莫愁 陆汉芝 李艳琴 秦会斌


摘要:為探讨退化喀斯特森林中不同植物叶片气孔导度对环境因子的响应特征及其适应性机制,以中国科学院普定喀斯特生态系统观测研究站沙湾监测研究区内的退化喀斯特森林为例,从中选取8种常见植物为研究对象,分别对其叶片气孔导度及气温、空气湿度、光照强度等环境因子进行测定分析。结果表明:(1)不同植物叶片气孔导度值日间主要在0.011~0.215 mol/(m2·s)之间变化。植物叶片气孔导度在同一时刻不同种间存在显著差异(P<0.05),而除构树[Broussonetia papyrifera (Linn.) L′Hér. ex Vent.]外,同种植物在不同时刻的差异均不显著(P>0.05)。(2)梓树(Catalpa ovata G. Don)的叶片气孔导度与光照强度呈显著中度负相关(r=-0.534,P=0.040),而马桑(Coriaria nepalensis Wall.)的则呈显著中度正相关(r=0.538,P=0.038)。构树的叶片气孔导度与相对湿度呈极显著高度负相关(r=-0.728,P=0.002),而与气温呈显著中度正相关(r=0.602,P=0.017)。厚果崖豆藤(Millettia pachycarpa Benth.)的叶片气孔导度与相对湿度呈显著中度负相关(r=-0.554,P=0.032)。(3)树种和测定时间对植物叶片气孔导度的交互作用不显著(P>0.05)。(4)研究区大部分植物对生境恶劣的喀斯特环境均具有较强的适应能力,叶片气孔导度的日间变化主要受植物自身遗传因素的控制,对外部环境条件改变的响应不敏感。
关键词:退化喀斯特森林;植物叶片;气孔导度;变异特征;环境响应
中图分类号:S718.45:Q948.11文献标识号:A文章编号:1001-4942(2020)04-0032-06
Abstract Stomatal conductance is one of the most direct gas exchange parameters that characterize plant photosynthesis. The response and adaptation of stomatal conductance of different plant leaves to environmental factors in degraded karst forest were studied in this article. The degraded karst forest located in Shawan monitoring area, the Karst Ecosystem Observation Research Station in Puding, Chinese Academy of Sciences was used as the experimental object. The stomatal conductance of eight common plants from the degraded karst forest, and the air temperature, air humidity, light intensity and other environmental factors were measured and analyzed. The results indicated that: (1) the stomatal conductance of the selected common plants mainly ranged from 0.011 to 0.215 mol/(m2·s) during daytime. There were significant differences in stomatal conductance among different species at the same time (P<0.05). However, the differences of the same species at different time were not significant (P<0.05) except Broussonetia papyrifera (Linn) L′Hér. ex Vent. (2) The correlation between stomatal conductance and light intensity was negatively moderate (r=-0.534, P=0.040) in Catalpa ovata G. Don, while that was positively moderate (r=0.538,P=0.038) in Coriaria nepalensis Wall. The stomatal conductance of Broussonetia papyrifera (Linn.) L′Hér. ex Vent. was very significantly negative correlated with relative humidity (r=–0.534, P=0.040), but significantly positive correlated with air temperature (r=0.602,P=0.017). The stomatal conductance of Millettia pachycarpa Benth. was moderately negatively correlated with relative humidity (r=-0.554,P=0.032). (3) The interaction of tree species and determination time on stomatal conductance was not significant (P>0.05). (4) Most of the plants in the research area have stronger adaptability to the harsh karst environment. The daytime variation of leaf stomatal conductance was mainly controlled by genetic factors, so it was not sensitive to the change of external environmental conditions.
Keywords Degraded karst forest; Plant leaves; Stomatal conductance; Variation characteristics; Response to environment
气孔是陆生植物叶片的一个重要结构,是高等植物与大气进行水气交换的主要通道,光合作用需要的CO2和蒸腾作用释放的水蒸汽均通过气孔进出植物体[1-4]。它随着环境的变化而变化,在植物中起着平衡调节的作用,是调控土壤-植被-大气连续体之间物质和能量交换的关键环节[5-7]。气孔关闭会阻碍外界 CO2进入叶片,限制光合作用暗反应速率,从而降低光合速率[8,9]。因此,气孔的开闭状态在一定程度上反映植物体内的代谢状况,并且气孔开闭的灵敏度也是植物十分重要的抗旱特征之一[3,10]。气孔导度是反映气孔行为最重要的生理指标,也是衡量植物和大气间水分、能量及 CO2平衡和循环的重要指标,探究气孔运动对环境因子的响应对于阐明气孔对光合作用及蒸腾作用的影响具有重要意义[2,3,6]。
退化喀斯特森林是喀斯特地貌上结构和功能退化并失去固有平衡的森林生态系统[11-13]。目前,利用喀斯特地区原生及适生能力强的植物来修复改善喀斯特脆弱生境是退化喀斯特森林生态系统修复重建的主要策略之一[14]。本研究以中国科学院普定喀斯特生态系统观测研究站沙湾监测研究区内的退化喀斯特森林中8种常见植物为研究对象,对其叶片气孔导度以及气温、空气湿度、光照强度等环境因子进行测定分析,旨在探讨不同植物叶片气孔导度对环境因子的响应及适应性机制,并为喀斯特森林生态系统的稳定性维护和退化生态系统修复重建中的植物选种和育种提供参考。
1 材料与方法
1.1 研究区概况
中国科学院普定喀斯特生态系统观测研究站沙湾监测研究区(北纬26°22′01″~26°22′08″,东经105°45′05″~105°45′09″)位于贵州省安顺市普定县城北约5 km处的城关镇陇嘎村沙湾组境内。该研究区下伏基岩为三叠系关岭组二段灰岩,土壤以石灰土为主,土层浅薄、分布不连续,质地为壤土;属亚热带季风湿润气候,多年平均气温16.0℃,年总积温5 829℃,年辐射量413.9 kJ/cm2,年降水量1 433 mm,无霜期340 d,雨热同季,降雨集中在5—9月;植被类型以藤刺灌丛为主,兼有次生常绿落叶阔叶林呈斑点状分布,主要物种有火棘[Pyracantha fortuneana (Maxim.) Li]、小果蔷薇(Rosa cymosa Tratt.)、粉枝莓(
Rubus biflorus Buch.-Ham. ex Smith)、槲蕨(Drynaria roosii Nakaike)、野桐(Mallotus tenuifolius Pax)、烟管荚蒾(Viburnum utile Hemsl.)、厚果崖豆藤(Millettia pachycarpa Benth.)、石岩枫[Mallotus repandus (Willd.) Muell. Arg.]、构树[Broussonetia papyrifera (Linn.) L′Hér. ex Vent.]、马桑(Coriaria nepalensis Wall.)、木通[Akebia quinata(Houttuyn)Decaisne]、樸树(Celtis sinensis Pers.)、铁线莲(Clematis florida Thunb.)、梓树(Catalpa ovata G.Don)等。
1.2 研究对象
于研究区内退化喀斯特森林中划定200 m2样方作为试验样地,从中选择长势一致且生长良好的8种常见植物:厚果崖豆藤、石岩枫、构树、马桑、木通、朴树、铁线莲和梓树为研究对象。其中,厚果崖豆藤是豆科(Leguminosae)崖豆藤属(Millettia)常绿藤本,石岩枫是大戟科(Euphorbiaceae)野桐属(Mallotus)常绿灌木,构树是桑科(Moraceae) 构属(Broussonetia)落叶乔木,马桑是马桑科(Coriariaceae)马桑属(Coriaria)常绿灌木,木通是木通科(Lardizabalaceae)木通属(Akebia)落叶藤本,朴树是大麻科(Cannabaceae)朴属(Celtis)落叶乔木,铁线莲是毛茛科(Ranunculaceae)铁线莲属(Clematis)落叶藤本,梓树是紫葳科(Bignoniaceae)梓属(Catalpa)落叶乔木。
1.3 试验设计
每种常见植物选择3株,在其树冠外围中上部,选择春梢上自顶端往下数第三片色泽正常、无病虫害且完全展开的成熟健康叶片作为测量对象。测前用蒸馏水淋洗干净,并挂牌标记。测定时,保持叶片自然生长角度不变,同一叶片重复测量5次取平均值。
1.4 测定指标及方法
于2019年9月(植物生长季)28日(晴天,无风)9—17时,采用LI-6400XT便携式光合测定系统分析仪(美国LI-COR公司产品)测定叶片气孔导度[Gs,mol/(m2·s),以H2O计],间隔2 h测定1批次,共测定5批次。测定时叶室内部光强为(1 078.3±99.1) μmol/(m2·s),由LI-6400-02B型红蓝光源提供;样品室气温为(32.6±2.0)℃,相对湿度为(26.2±6.3) %,CO2浓度为(394.9±8.5)μmol/mol,参比室CO2浓度为(403.5±7.9) μmol/mol,均由LI-6400便携式光合测定系统分析仪测定提供。
植物叶片周围气温、相对湿度均采用DT-321S型温湿度测量仪(CEM深圳华盛昌机械实业有限公司产品)测定,光照强度采用AS823分体式照度计(香港希玛仪表集团有限公司产品)测定。该类指标均在距离气孔导度测定所用叶片1 cm2范围内,紧随气孔导度测定完成后测定。
1.5 数据分析
利用Microsoft Excel 2010软件整理数据及作图,IBM SPSS Statistics 19.0软件进行统计分析。统计分析包括描述性统计分析、单因素方差分析和多因素方差分析及相关分析等。
2 结果与分析
2.1 植物叶片气孔导度的日变化特征
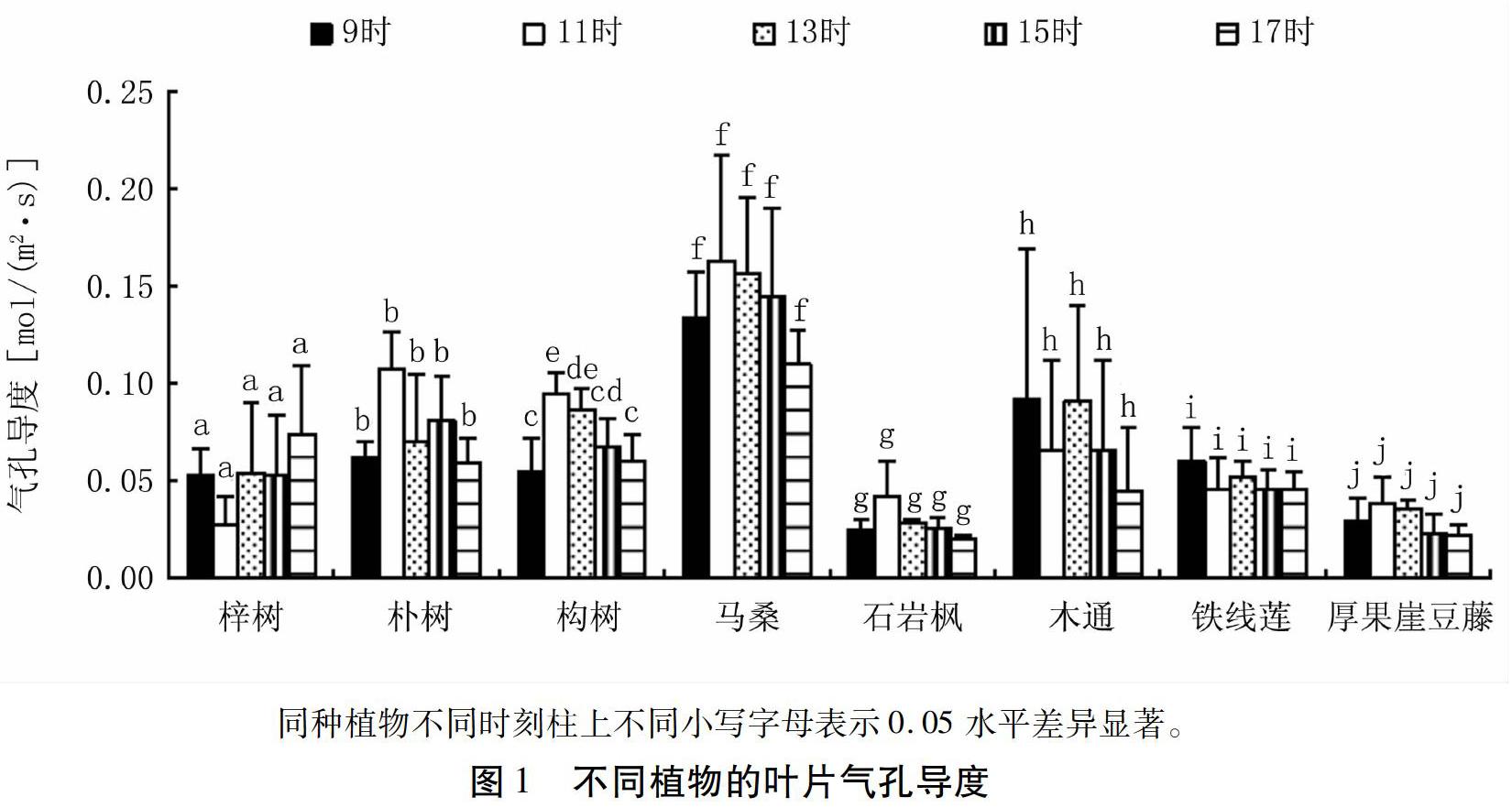
由表1可以看出,不同植物叶片气孔导度值在0.011~0.215 mol/(m2·s)之间,平均值从大到小依次为马桑>朴树>构树>木通>梓树>铁线莲>厚果崖豆藤>石岩枫。气孔导度最大值与最小值之比和变异系数均较小,说明这些植物叶片气孔导度在日间波动不大。构树的偏度值为负数,呈负偏态分布,其余7种植物均呈正偏态分布。
由表2可以看出,不同时刻叶片气孔导度平均值从大到小依次为11时>13时>9时、15时>17时。同一时刻的最大值与最小值之比和变异系数相对同种植物日变化的最大值与最小值之比和变异系数(表 1)均较大,表明同一时刻不同植物间的差异要大于同种植物不同时刻的差异。
由图1可以看出,研究区8种常见植物叶片气孔导度值的日变化中,落叶藤本植物木通和铁线莲的变化趋势一致,数值均以9时最大,17时最小;常绿灌木马桑和石岩枫的变化趋势一致,均在11时达到最大值,17时降到最低。常绿藤本植物厚果崖豆藤和落叶乔木朴树的叶片气孔导度值也在11时达到最大,17时降到最低。落叶乔木中,构树叶片气孔导度的最大值和最小值均出现在上午,以11时最大,9时最小,而梓树的最大值却出现在17时,最小值出现在11时。说明不同植物叶片气孔导度日变化趋势并不完全相同。
方差分析和多重比较结果表明,木通、铁线莲、厚果崖豆藤、马桑、梓树、石岩枫、朴树的叶片气孔导度值在日间各时刻的差异均不显著(P>0.05),而构树的叶片气孔导度值在11时与9时(P=0.005)、11时与15时(P=0.035)、11时与17时(P=0.010)、13时与9时(P=0.015)、13时与17时(P=0.032)均存在显著差异。说明研究区8种常见植物中,除构树叶片气孔导度随时间的改变发生显著变化外,其它植物叶片气孔导度日间均无显著变化。
2.2 植物叶片气孔导度的种间差异特征
由表3可以看出,9时,不同植物的叶片气孔导度差异显著(P=0.011),多重比较后按同类子集可将其分为石岩枫+厚果崖豆藤+梓树+构树+铁线莲+朴树[JP](P=0.202)、梓树+构树+铁线莲+朴树+木通(P=0.180)、木通+马桑(P=0.119)3类。11时,叶片气孔导度的种间差异显著(P=0.000),多重比较后按同类子集可将其分为梓树+厚果崖豆藤+石岩枫+铁线莲+木通(P=0.163)、石岩枫+铁线莲+木通+构树(P=0.053)、木通+构树+朴树(P=0.106)、马桑(P=1.000)4类。13时,种间气孔导度差异显著(P=0.001),[JP]多重比较后按同类子集可将其分为石岩枫+厚果崖豆藤+铁线莲+梓树+朴树(P=0.137)、厚果崖豆藤+铁线莲+梓树+朴树+构树+木通(P=0.054)、马桑(P=1.000)3类。15时,种间气孔导度差异显著(P=0.002),多重比较后按同类子集可将其分为厚果崖豆藤+石岩枫+铁线莲+梓树+木通+构树(P=0.094)、铁线莲+梓树+木通+构树+朴树(P=0.171)、马桑(P=1.000)3类。17时,种间气孔导度差异显著(P=0.001),多重比较后按同类子集可将其分为石岩枫+厚果崖豆藤+木通+铁线莲(P=0.162)、厚果崖豆藤+木通+铁线莲+朴树+构树(P=0.052)、木通+铁线莲+朴树+构树+梓树(P=0.133)、马桑(P=1.000)4类。
2.3 植物叶片气孔导度与环境因子的相关性
两变量相关系数(r)可反映其相关性强弱。一般来说,|r|在0.00~0.33之间为低度相关,0.33~0.67之间为中度相关,0.67~1.00之间为高度相关[15]。由表4可以看出,不同植物叶片气孔导度对环境因子的响应程度不同。梓树的叶片气孔导度与光照强度呈显著中度负相关(r=-0.534,P=0.040),而马桑的则呈显著中度正相关(r=0.538,P=0.038)。构树的叶片气孔导度与相对湿度呈极显著高度负相关(r=-0.728,P=0.002),而与气温呈显著中度正相关(r=0.602,P=0.017)。厚果崖豆藤的叶片气孔导度与相对湿度呈显著中度负相关(r=-0.554,P=0.032)。
2.4 植物自身遗传因素和测定时间对叶片气孔导度的交互作用
为了解研究区植物自身遗传因素和环境因子对叶片气孔导度的交互影响情况,以植物种和测定时间为控制变量、植物叶片气孔导度为观测变量建立固定效应的饱和模型进行多因素方差分析,结果(表5)表明,研究区的植物种和测定时间对植物叶片气孔导度的影响没有产生显著的交互作用(P>0.05);植物叶片气孔导度在不同种间存在极显著差异(P=0.000),而在不同时刻的差异不显著(P=0.124),表明物种是影响植物叶片气孔导度的主要因素。该结论与前述单因素方差分析、相关分析和多重比较结果一致。
3 讨论与结论
植物叶片的光合生理特征既受自身遗传因素影响,也与外部环境条件有关[16-19]。一日之中,太阳辐射随时间变化而变化,这可能引起各种生态因子发生改变,使得同一生境植物在不同时间的環境条件有可能不同[20,21]。气孔导度是表征植物光合作用最直接的气体交换参数之一,较高的气孔导度对应于更高的光合速率[13,22]。上午植物叶片的气孔逐渐张开,气孔导度逐渐增大以提高光合速率,但气孔导度并非随光强的增加一直增加[23],当遭遇干旱或高温胁迫时,为减少蒸腾损失气孔会逐渐关闭,气孔导度降低使得胞间CO2浓度不足,光合作用可能会受到抑制[24]。
