石榴毡蚧种群的空间格局及其抽样模型的建立
2020-07-01孙丹萍王朝阳
徐 睿,孙丹萍,王朝阳
(河南林业职业学院,河南 洛阳 471002)
在自然界中,任何昆虫的种群都是以一定的空间格局进行分布的。种群空间格局是指昆虫种群在空间上的分布方式[1]。空间格局是昆虫种群的重要特性,也是昆虫的生物学特性,代表着昆虫生存空间需求的特征及动态变化,反映了昆虫在生态环境中的生存方式,并受到了外界环境条件的影响[2-4]。了解昆虫的种群空间格局对掌握种群的发展、扩散、猖獗、消亡的预测预报及制订可持续害虫控制策略等有重要的意义[2,5]。昆虫的种群空间格局受到许多因素的影响,包括生态环境、昆虫习性等。研究种群空间格局的生态学目的之一是了解环境对种群的影响,不仅要分析影响其分布的环境因素,还要研究环境对种群分布产生的影响[6]。
有关昆虫种群空间格局抽样模型的研究报道较多,抽样模型必须符合4 个特点:准确、简单、迅速、廉价[2]。目前,比较普遍采用的抽样模型包括聚集度指标法、线性回归法、扩散型指数法等[7-11]。常亚文等[7]研究了三叶斑潜蝇幼虫在番茄和豇豆上的空间格局分布,同时对抽样技术进行了研究。结果表明,三叶斑潜蝇幼虫在2 个寄主上均呈聚集分布,且以个体群形式存在。同时,采用Kc 法、Iwao 法及Taylor 幂法计算出了在不同精度下三叶斑潜蝇田间的理论抽样数。陈红星等[12]通过对垂叶榕不同方位和高度的随机取样,运用5 种聚集度指标以及Taylor 幂法和Iwao 回归模型分析和测定了榕管蓟马Gyuaikothrips uzedli的种群空间格局,同时应用Iwao 理论抽样模型确定了榕管蓟马不同种群密度与不同精度要求下的理论抽样数。潘杰[2]采用地统计学方法分析了受害程度不同的油松纯林和混交林林分内红脂大小蠹种群的空间分布格局,发现轻度受害纯林与重度受害纯林林分内红脂大小蠹种群均表现为较强的空间聚集性,而混交林受害林分内其种群表现为随机分布。受害程度不同的纯林林分内红脂大小蠹种群空间格局具有一定的差异,而且纯林与混交林林分内其种群空间格局也有较大的差别。近年来,随着虫害防控技术的升级以及抽样技术的成熟,我国在森林保护领域的害虫控制、预测预报模型等方面的研究均获得了较好的成果。
石榴Punica granatumL.的市场价值高,种植前景好,在我国种植范围广。软籽石榴新品种是目前我国大力发展的经济树种之一,种植面积不断扩大,经济效益大幅提升。但近年来,石榴毡蚧的危害呈现日益加重的趋势。石榴毡蚧又名紫薇绒蚧、石榴粉蚧、石榴绒蚧,属同翅目Homoptera 绒蚧科Eriococcidae 绒粉蚧属Eriococcus,为害石榴、苹果、紫薇、大豆等多种花果树木和农田作物,其若虫和雌成虫寄生于小枝、叶片主脉基部、芽腋、嫩梢或枝干等部位刺吸汁液,常造成树势衰弱,生长不良。虫口密度大时,枝叶发黑,叶片早落,开花异常,严重影响树木生长和果品的质量、产量及商品价值。2013年,石榴毡蚧被列入我国林业危险性有害生物名单[13-15]。昆虫的空间分布状况是其种群的重要生态学属性[16]。通过研究石榴毡蚧种群的空间分布格局,了解其空间结构,从而确定田间不同允许误差下的最大抽样数量及序贯抽样技术,使虫情调查结果能够更加客观地反映种群的动态变化趋势,及时掌握最有利防治时机,对石榴毡蚧的科学测报和高效防控具有重要的指导 意义[17-21]。
1 材料与方法
1.1 取样方法
2016年7—8月,石榴毡蚧发生期,在郑州市和灵宝市的果园、绿地等不同立地条件下,选择8个样地,每个样地按照平行线法选取10 株树,每株树分别按东、西、南、北、中5 个方位,每方位选取4 片叶或20 cm 小枝1 根,检查统计每个处理的蚧虫数量。
1.2 空间分布型测定
1.2.1 聚集度指标法
根据调查数据,以样地为单位,计算样本数均值m(头/叶)和样本方差V,采用以下聚集度指标[2-9]进行检验分析。
1)Caasier 扩散系数(C值)
C=V/m。
当C=1 时,种群为随机分布;当C>1 时,为聚集分布;当C<1 时,为均匀分布。
2)Waters 负二项分布参数(k值)
k=m2/(V-m)。
k值愈小,种群聚集度愈大;当k>8 时则接近Poisson 分布。
3)Lloyd 聚块性指标(m*/m)
m*=m+V/m- 1。
当m*/m=1 时,种群为随机分布;当m*/m>1时,为聚集分布;当m*/m<1 时,为均匀分布。
4)Cassie & Kunou 指标(CA)
CA= (V-m)/m2。
当CA=0 时,种群为随机分布;当CA>0 时,为聚集分布;当CA<0 时,为均匀分布。
5)David & Moore 丛生指标(IDM)
IDM=V/m- 1。
当IDM=0 时,种群为随机分布;当IDM>0 时,为聚集分布;当IDM<0 时,为均匀分布。
6)兰星平Lα指标
Lα=m-m/V+ 1。
当Lα/m=1 时,种群为随机分布;当Lα/m>1时,为聚集分布;当Lα/m<1 时,为均匀分布。
7)张连翔Z/V指标
Z=V/m- 1+V。
当Z/V=1 时,种群为随机分布;当Z/V>1 时,为聚集分布;当Z/V<1 时,为均匀分布。
1.2.2 线性回归分析法
1)TaylorV-m幂法则(V=amb)回归模型
lgV= lga+blgm。
式中:V为样本方差,m为样本平均值,a、b为引入的参数。当lga=0 且b=1 时,种群为随机分布;当lga>0 且b=1 时,种群为聚集分布;当lga>0且b>1时,种群为聚集分布;当lga<0且b=1时,种群为均匀分布。
2)Iwaom*/m回归模型
m*=a+bm。
式中:m为样本均值,a、b为引入参数。当a=0且b=1 时,种群为随机分布;当a>0 且b=1 时,为聚集分布,分布的基本成分为随机分布的个体群;当a=0 且b>1 时为负二项分布;当a>0且b>1 时为聚集分布的个体群。当a=0 且b<1或0 >a>-1 且b=1 时为均匀分布。
3)兰星平c′-m模型
c′=a+bm。
式中:a、b为待估参数,m为均值。当c′、m线性相关显著时,a、b不同组合型揭示种群不同的空间分布型。
c′ = 1 /c+m。
式中:c为扩散系数。当a≤1 且b<1 或a<1且b≤1 时,为聚集分布;当a=1 且b=1 时,为随机分布;当a≥1 且b>1 或a>1 且b≥1 时,为均匀分布。
1.3 聚集因素分析
运用Blackith 种群聚集均数(λ)检验石榴毡蚧种群的聚集原因。

式中:k为负二项分布的指数,γ是χ2分布表中自由度等于2k概率(P=0.5)对应的χ2值。在此基础上进行λ-m线性回归分析,确定聚集原因。当λ<2时,种群聚集原因可能是某些环境因子的作用;当λ≥2 时,种群聚集原因是环境因子和昆虫习性的综合作用。
1.4 抽样分析
1.4.1 理论抽样分析
在确保调查质量的前提下,须抽取的最少样本数为理论抽样数。根据Iwaom*/m回归模型提出的理论抽样模型公式进行分析确定。
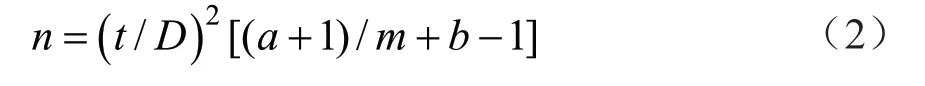
式中:t为置信度,实际调查中t取1;D为允许相对误差;m为估计的虫口密度;a、b为Iwaom*/m回归模型参数。
1.4.2 序贯抽样分析
用Iwaom*/m序贯抽样决策模型公式,通过拒绝限和接受限(二者合称“决策限”)来确定石榴毡蚧的序贯抽样模型公式及最大理论抽样数公式。
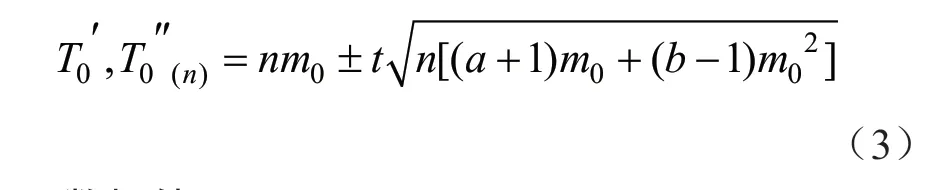
1.5 数据处理
调查数据采用DPS 软件进行处理分析。
2 结果与分析
2.1 种群空间分布型
2.1.1 聚集度指标检验
应用7 种聚集度指标分别对调查数据进行检验分析,结果见表1。由表1 可知,扩散系数C和聚集指标m*/m值均大于1;负二项分布k值,2 个样地k<1,6 个样地k>1,且均小于8;Cassie 指标CA和丛生指标IDM值均大于0;Lα指标Lα/m和Z指标Z/V值均大于1。这些聚集度指标数值均符合聚集分布的检验标准,表明石榴毡蚧种群的空间分布型为聚集分布。
2.1.2 回归模型检验
石榴毡蚧种群空间分布的回归模型分析结果见表2。由表2 可知,根据Taylor 幂法则回归模型检验结果,lga=0.696 2 >0,b=1.501 5 >1,种群为聚集分布,分布具有密度依赖性,说明石榴毡蚧种群在任意密度下都是聚集的,其聚集强度随种群密度升高而增加。根据Iwaom*/m模型检验结果,a=14.886 0 >0,说明石榴毡蚧个体间相互吸引,分布的基本成分是个体群;b=1.382 6 >1,说明种群个体群为聚集分布,且符合负二项分布。根据兰星平c′-m模型检验结果,a=0.058 4 <1,b=0.999 3 <1,说明石榴毡蚧种群为聚集分布。 3 种模型拟合优度均在0.8 以上,拟合效果较好,表明石榴毡蚧种群的空间分布型为聚集分布,这与采用聚集度指标法测定的结果相一致。
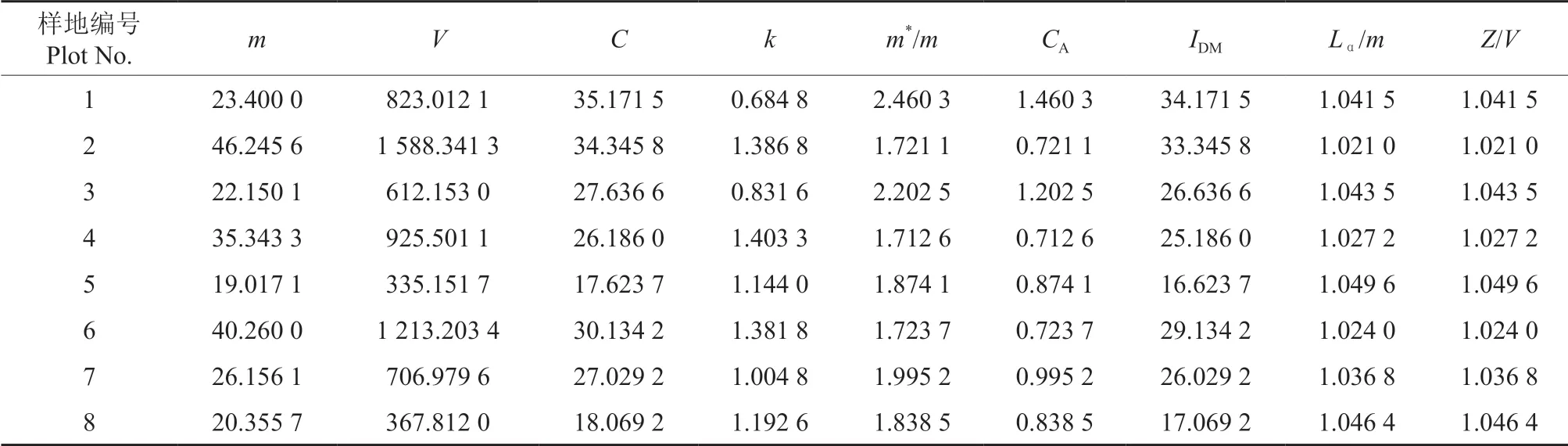
表1 石榴毡蚧种群的聚集度指标分析结果Table 1 Analysis result of aggregation degree indexes of E.lagerstroemiae population

表2 石榴毡蚧种群空间分布的回归模型分析结果Table 2 Analysis result of spatial distribution regression models of E.lagerstroemiae population
2.1.3 聚集因素分析
通过式(1)计算Blackith 种群聚集均数(λ),结果见表3。由表3 可知,λ值均远大于2,说明其聚集可能是由于某些环境因子作用所引起,也可能是昆虫自身聚集习性作用的结果。
对各组样本数均值(m)与聚集均数(λ)进行相关分析,得出回归方程:λ=0.509 7m-0.342 8,R2=0.723 7(图1)。由图1 可以看出,石榴毡蚧聚集均数随平均虫口密度增加而不断增大。若令λ=2,则m=4.596 4。即当每叶石榴毡蚧虫口数量小于4.596 4 头时,其聚集原因是由环境因子引起;当每叶虫口数量大于或等于4.596 4 头时,其聚集可能是由环境因子引起,也可能是由石榴毡蚧自身的聚集习性引起。本研究中,每叶石榴毡蚧虫口数量均大于4.596 4 头,表明石榴毡蚧聚集是自身的生活习性、行为及环境因子综合影响的结果。经调查发现,自然情况下石榴毡蚧大多营固定生活,活动范围小,自主扩散能力弱,分布具有局限性,个体间相互吸引,形成基本个体群。石榴毡蚧繁殖能力强,随着繁殖量的增加,个体数量和虫口密度不断增加,个体群规模也随之增大。此外,石榴毡蚧在果园、绿地等多呈团块状分布,在一定程度上限定了其扩散的空间范围,从而呈现团块状密集分布的特点。这些因素与数据分析结果基本吻合,说明石榴毡蚧聚集是其自身的聚集习性与环境因子综合影响的结果。
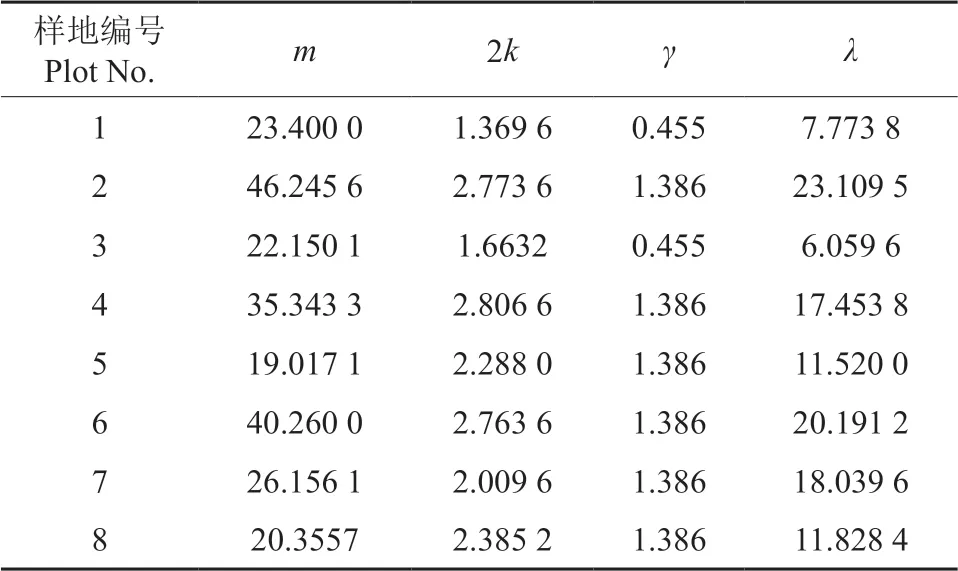
表3 石榴毡蚧种群聚集均数与样本数均值分析结果 Table 3 Analysis result of average aggregation sizes and average sample number of E.lagerstroemiae population
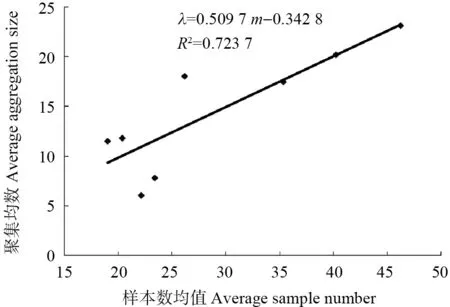
图1 石榴毡蚧种群聚集均数(λ)与样本数均值(m)的 关系Fig.1 Relation between average aggregation size (λ) and average sample number (m) of E.lagerstroemiae population
2.2 抽样模型的建立
2.2.1 理论抽样模型的建立
根据Iwao 理论抽样公式,将已经建立的Iwaom*/m回归模型中的参数a=14.886 0、b=1.382 6、t=1,代入式(2),可建立石榴毡蚧的最适理论抽样模型n=(1/D)2(15.886 0/m+ 0.382 6),确定不同虫口密度、不同误差条件下的最适理论抽样数(表4)。
由于寄主植物数量常是有限的,即总体有限,故在实际抽样中应校正公式:
n′=n/(1 +n/N)。
式中:N为总体数,n′为校正后的理论抽样数。
2.2.2 序贯抽样模型的建立
根据蚧虫一般防治指标为5 头/叶,将t=1、m0=5、a=14.886 0、b=1.382 6代入式(3),可得石榴毡蚧种群序贯抽样决策模型公式:若取一系列n值(n=5,10,15,20,…),可得不同累计虫口数相应抽样数的上、下限值(T′0(n)、T″0(n)),结果见表5。
在实际应用中,若抽取样本的累计虫口数总是介于上、下限(T0′、T0″)之间,则实际抽取样本数应不大于最大理论抽样数nmax,即nmax=[(14.886 0+1)/m0+1.382 6-1]/D2。 令D=0.1,则nmax=356;D=0.2,则nmax=89,并以其最接近的界限方程得出结论。
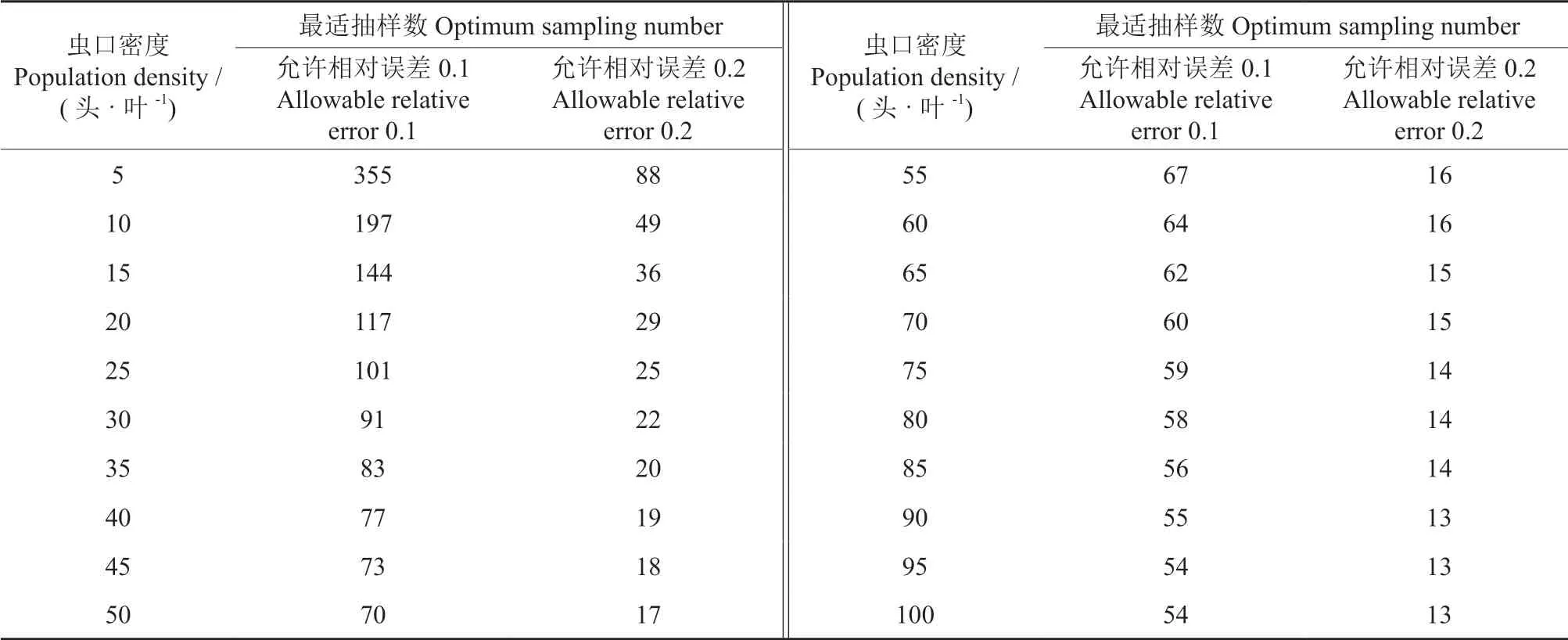
表4 石榴毡蚧种群最适理论抽样数Table 4 Optimum theoretical sampling number of E.lagerstroemiae population
3 结论与讨论
本研究结果表明,石榴毡蚧种群空间分布型为聚集型负二项分布,且分布具有密度依赖性,石榴毡蚧种群在任意密度下都是聚集的,其聚集强度随种群密度升高而增加。石榴毡蚧种群聚集分布是由其生活习性及环境因素共同影响的结果。总的来说,这是石榴毡蚧种群对生态环境适应的结果。

表5 石榴毡蚧种群序贯抽样数Table 5 Sequential sampling number of E.lagerstroemiae population
在进行田间调查时,为了获得准确的调查资料,可结合生产实际,应用最适理论抽样数模型和最佳序贯抽样模型进行序贯抽样。当调查的累计虫口数量达到预定虫口数量指标时,应停止调查,累计虫口数量除以取样数,即平均虫口密度。在生产实际中,据此可以精准监测虫情,把握最有利防治时机,制定经济、安全、高效的防控对策,控制石榴毡蚧的危害。
3.1 石榴毡蚧种群空间格局
分析种群空间格局是种群生态学的重要研究内容,可揭示种群在生境空间的扩展情况[22]。目前,有关蚧类种群空间格局的研究主要集中在扶桑绵粉蚧Phenacoccus solenopsis、松突圆 蚧Hemiberlesia pitysophila、矢尖蚧Unaspis yanoncnsis、杨盾蚧Diaspidiotus slavonicus等[22-25]。 有关石榴毡蚧种群空间格局的研究鲜见报道。昆虫种群的空间格局是认识昆虫的生态学特性的基础之一。本研究中发现,石榴毡蚧种群空间分布为聚集型分布,这与黄俊等[22]得出的检疫性有害生物扶桑绵粉蚧雌成虫在大花马齿苋上的空间格局是相似的,且两者均具有密度依赖性。侯沁 文[23]应用聚集度指标和回归分析,对松突圆蚧空间分布型进行研究,发现松突圆蚧的若虫在松林中任意密度下均呈聚集分布,聚集度随虫口密度增大而增强,成虫在松林中呈均匀分布。这与石榴毡蚧在任意密度下都是聚集的,且聚集强度随种群密度升高而增加的结论相似。江西林等[24]研究了矢尖蚧在天竺桂叶片上的空间格局,发现其为聚集型分布,这与石榴毡蚧种群的空间格局是一 致的。
3.2 石榴毡蚧种群空间分布的影响因素
影响昆虫种群空间分布的因素包括昆虫的生活习性和外界环境。石榴毡蚧Blackith 种群聚集均数(λ)均远大于2,由此可知种群聚集原因是环境因素和昆虫习性共同作用的结果。一方面,石榴毡蚧生命力和繁殖力极强,尤其是在春末秋初,温湿条件较适宜其发生和繁殖,繁殖速度快,短期内虫口密度常会急剧增加;另一方面,石榴毡蚧种群相对固定的生活方式和雌成虫固定产卵的习性是形成其聚集型分布格局的主要原因,具体原因有待进一步深入研究。对于松突圆蚧而言,若虫聚集分布的原因主要与其集中产卵的方式有关。成虫集中产卵后,其中大部分若虫就近孵化,大量聚集孵化后由于食物竞争的原因,大量若虫发育迟缓或死亡,成活率较低,小部分若虫爬到远处,因分散而营养充足,发育成成虫,成活率较高,则成均匀分布[23]。天竺桂叶片上矢尖蚧的空间格局为聚集分布,可能与初孵若蚧的爬行行为和矢尖蚧天敌的聚集行为有关[24]。由此可知,对于这些昆虫而言,影响种群空间分布的主导因素是其生活习性。此外,石榴等寄主植物的栽培方式及特点可能是其形成聚集型空间分布的重要因素之一,生态环境中气候条件也直接影响着石榴毡蚧种群的分布状况。由此说明,石榴毡蚧种群空间分布与寄主植物栽培配置方式、树木生长状况、气候条件及虫口密度等因素密切相关。
3.3 石榴毡蚧种群抽样模型研究
确立最适抽样数和抽样模型是种群数量预测预报的基础。目前,通常是通过预估抽样模型的种群参数来确定虫害的发生情况。贾蕊娟等[25]应用分布型指数研究了凤凰木夜蛾Pericyma cruegeri幼虫在凤凰木Delonix regia上的空间分布型,利用Iwao 模型、Taylor 幂法则分析聚集原因,同时确定了幼虫的最适理论抽样数及序贯抽样表,与本研究中所采用的抽样模型类型一致,都能准确反映昆虫的种群情况。此外,付园园等[26]在传统抽样分析方法的基础上引入地统计学的方法,对不同林分类型的杨树林内杨盾蚧的分布进行半变异函数拟合,建立高斯模型、指数模型、线性模型等。另外,传统的方法存在一定的弊端,当混交林虫情指数过低时,存在不能建立回归方程、难以确认样地之间空间分布的差异、无法描述虫情峰值大小等问题,引入地统计学的方法后,通过制作昆虫分布图,可以形象地描述昆虫分布情况,取得更好的效果。因此,为了更加清晰地描述石榴毡蚧在不同空间结构的格局,可以尝试引入地统计学的方法来研究石榴毡蚧的防治[26-28]。
