冻融循环条件下水分和氮添加对黑土有机碳矿化的影响
2020-07-01郭晓丽戴闪闪李禄军
郭晓丽,何 朋,戴闪闪,李禄军
(1.中国科学院 东北地理与农业生态研究所,黑龙江海伦农田生态系统国家野外科学观测研究站,黑龙江 哈尔滨 150081;2.中国科学院大学,北京 100049)
0 引 言
作为陆地上第一大有机碳库,土壤有机质对土壤肥力以及全球碳循环具有重要影响。土壤有机碳矿化是影响土壤碳库大小和土壤碳循环的一个重要过程,影响着土壤中营养元素的释放和供应、温室气体的排放以及土壤质量的保持等。土壤有机碳的矿化量和速率受到土壤理化性质和外界环境因子的共同影响,其中,土壤水分和土壤养分是影响土壤有机碳矿化的重要因素[1-4]。
Fang和Moncrieff[5]在室内培养时发现,在20%~50%体积含水量条件下,水分对土壤呼吸的影响不显著。Devevre和Horwath[6]发现,非淹水状态下土壤有机碳和外源添加物的矿化速率均高于淹水状态,淹水使土壤有机碳的矿化量降低,Shaffer等[7]认为,旱地土壤有机碳的分解速率高于湿地主要是因为旱地土壤含氧量更高。然而,黄东迈等[8]应用14C示踪技术得到了与之前观点相反的研究结果,发现在淹水土壤中的有机碳分解速率高于旱地土壤,黄耀[3]也发现,淹水条件下土壤有机碳的矿化量最大。李忠佩等[9]认为淹水提高了土壤有机碳矿化量的主要原因是淹水增加了可溶性碳的含量。因此,水分变化对土壤有机碳矿化的影响存在很大的不确定性。
除了水分,氮素有效性是影响土壤有机碳矿化的另一重要因素。由于土壤碳循环和氮循环紧密耦合,氮素水平的改变可以显著影响土壤碳循环过程。有研究发现,氮素添加通过影响土壤微生物量和微生物对有机碳的利用效率,进而改变土壤有机碳矿化速率[10-11]。而且,在不同生态系统中,添加氮素对土壤有机碳矿化具有不同的影响,可表现为促进[12]、抑制[13]或无显著影响[14]。因此,氮素添加对土壤有机碳矿化的影响结果不一,主要是由于氮素添加会同时影响土壤有机碳矿化的多个直接或间接途径,从而对土壤有机碳矿化产生不一致的影响[15]。同时,土壤水分状况等因素的变化和氮素水平的变化对土壤有机碳矿化速率还会产生交互影响[16]。
冻融循环是土壤温度在0℃上下变化导致的土壤冻结和融化现象。在冻融循环过程中,土壤性质的变化本质上是由于土壤中水的相态在液态和固态之间的相互转变而引起的[17]。冻融循环作用能够改变土壤物理结构,影响土壤生物地球化学循环过程[18-19]。冻融作用通过改变土壤水分体积使土壤团聚体破碎,改变土壤物理性质,例如土壤团聚体稳定性、水热传导和水分入渗等过程[20],还会直接或间接影响相关土壤生物和化学性质,如低温导致土壤微生物死亡、团聚体的破碎促进土壤可溶性有机碳和氮的释放等[21]。因此,冻融循环条件下,土壤水分的变化会通过改变土壤物理、生物及化学过程来影响土壤有机碳矿化[22-23]。然而,受研究条件等诸多因素限制,有关冻融循环对黑土农田土壤有机碳矿化影响的研究较少,这限制了对黑土农田土壤有机碳变化规律的系统了解。
我国东北黑土地因其土壤有机质和养分含量高而闻名,但是,近年来黑土退化,土壤有机质剧烈下降,严重威胁黑土农田可持续生产能力[24]。研究黑土农田土壤有机碳矿化的影响因素及其变化规律对于土壤碳的固定具有重要意义。我国东北地处冷凉区域,在早春和晚秋季节该区域内存在强烈的冻融现象[25]。此外,受极端降水的影响,东北黑土农田在早春或晚秋季节土壤含水量变异较大,同时,长期大量的氮肥施用很容易造成农田土壤氮素的积累。土壤水分和氮素水平的变化可能会协同影响土壤有机碳矿化过程。加之该时期土壤冻融循环现象明显,使得土壤有机碳矿化对土壤水分和氮素水平的响应产生更大的不确定性。因此,本研究选取东北典型农田黑土为研究对象,采用室内模拟冻融循环的培养方法,研究在冻融循环条件下不同土壤含水量和氮添加对土壤有机碳矿化的影响,为揭示冻融期土壤有机质演变规律提供重要资料。
1 材料与方法
1.1 样品采集
本研究供试土样取自我国东北黑土区中心黑龙江海伦市(47°26′N,126°38′E),该区域属于温带大陆性季风气候,海拔高度为240 m,年平均气温为1.5 ℃,极端最低温度为-39.5 ℃,多年平均降雨量约为550 mm,无霜期125 d,冻结期长达7个月,每年10月中下旬至11月中旬和次年3月上旬至4月下旬存在频繁的冻融循环。
土壤样品取自农田土壤表层(0~20 cm),采样地为玉米(Zeamays)-大豆(Glycinemax)轮作制,取样时当季作物为大豆。2017年5月初,在春播施肥前随机多点采集土壤,均匀混合,去除肉眼可见的植物残茬和石块等杂物,新鲜土样过2 mm筛后分成两份,一部分土壤样品保存在4 ℃冰箱中,用来做室内培养实验,另一部分土壤样品自然风干后,用于测定土壤pH、有机碳和全氮等土壤基本性质(表1)。

表1 土壤基本性质(n=3)
注:表中数据为平均值±标准误差。
Note: Data represent mean±SE.
1.2 实验设计与测定方法
取过筛后的土壤,调节含水量至土壤田间持水量(WHC)的40%,在22℃的恒温培养箱中恒温预培养7 d,使土壤微生物活性和功能得以恢复。预培养结束后,称取相当于50 g干土的预培养土壤置于150 ml塑料培养瓶中,用于土壤有机碳矿化的培养实验。
本实验设置4个处理,分别为:60%WHC+不添加氮(LWLN)、80%WHC+不添加氮(HWLN)、60%WHC+添加氮(LWHN)和80%WHC+添加氮(HWHN),每个处理设置3个重复。另外,设置3个不加土壤的培养瓶作为空白。其中,60%WHC是土壤微生物活动的适宜含水量,80%WHC是高含水量处理;氮素以(NH4)2SO4溶液的形式添加,添加N含量为0.04 g·kg-1,添加依据研究区实际生产中田间氮素平均施用量的一半(70 kg·hm-2)。在调节土壤含水量时要把(NH4)2SO4溶液中所含水分计算在内。
依据相关文献[26-27],将冻结温度设置为-10 ℃,融化温度为10 ℃,冻结阶段和融化阶段的持续时间都为12 h,即24 h为一个冻融循环,共设置19个冻融循环周期。冻融循环过程中,冻结温度和融化温度均通过下述温度控制模块自动调控,实现增温和降温的连续进行。
土壤有机碳矿化由土壤有机质在矿化过程中CO2的排放量表征,土壤CO2排放速率采用全自动变温土壤微生物呼吸系统(PRI-8800,Pri-Eco,中国北京)测定。该培养系统由温度自动控制模块和CO2同位素分析仪(G2131-i,Picarro,美国)组成,可实现实时、连续和高频率地测量土壤CO2排放速率,每个样品测定时长3 min。在本研究中,土壤有机碳矿化速率(Rs)由固定时间段CO2浓度变化的斜率计算[28]:
Rs=C×V×α×β / m
式中,Rs为土壤有机碳矿化速率(ug·g-1·h-1);C为CO2浓度变化的斜率;V为培养瓶和气管的总体积(L);m为干土重(g);α为CO2质量转换系数;β为时间转换系数。
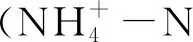
1.3 统计分析
实验数据采用Microsoft Excel 2013进行处理和计算,并通过SPSS 18.5使用双因素方差分析(two-way ANOVA)对不同处理间的差异显著性进行统计分析(P<0.05),由于土壤含量与氮添加对大多数指标都有显著交互影响,因此,进一步使用单因素方差分析(one-way ANOVA)对不同处理之间所有指标均值的差异进行显著性分析。采用Origin 8.5进行作图。
2 结果与分析
2.1 冻融条件下水分、氮添加对土壤无机氮和微生物量的影响
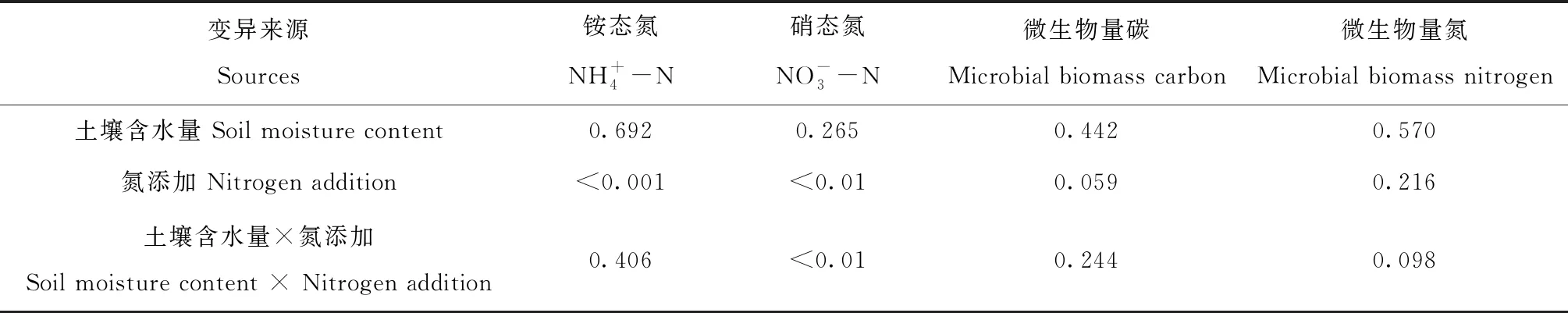
表2 土壤含水量和氮添加对土壤微生物量和无机氮影响的方差分析结果
冻融循环培养结束后,4个处理LWLN、HWLN、LWHN和HWHN的土壤MBC和MBN含量与冻融培养前相比均显著下降,土壤MBC分别降低了51.1%、39.2%、31.2%和33.8%(P<0.05,图2A),除HWHN处理外,土壤MBN分别下降了33.6%、61.2%和39.5%(P<0.05,图2B)。无论添加氮还是不添加氮,水分添加对土壤MBC和MBN含量均无显著影响(P>0.05,表2,图2)。在60%WHC土壤含水量条件下,氮添加显著增加了土壤MBC含量(P<0.05,图2A);在80%WHC土壤含水量条件下,氮添加处理显著增加了土壤MBN含量(P<0.05,图2B)。
2.2 冻融循环条件下不同土壤水分和氮添加对土壤有机碳矿化的影响
4个处理(LWLN、HWLN、LWHN和HWHN)在整个冻融循环期土壤有机碳累积矿化量的变化范围为664~1 930 ug·g-1,其中,融化期(10 ℃)和冻结期(-10 ℃)分别约占冻融循环期总矿化量的54%和46%。在融化期,氮添加主效应对土壤CO2累积排放量具有显著影响(P<0.01),而土壤含水量(P=0.08)及其与氮添加的交互作用(P=0.09)对土壤CO2累积排放量无显著影响(表3);在冻结期和整个冻融期,氮添加(P<0.01)及其与土壤含水量的交互作用(P<0.05)对土壤CO2累积排放量均有显著影响(表3)。
在融化期,HWLN处理的土壤CO2累积排放量显著低于LWLN处理(P<0.05),而在整个冻融期和冻结期的土壤CO2累积排放量在HWLN和LWLN处理之间的差异均不显著(图3);在添加氮的情况下,具有较高土壤含水量的土壤在冻结期CO2累积排放量显著高于较低土壤含水量(P<0.05),而不同水分处理对整个冻融期和融化期的土壤CO2累积排放量的影响不显著。
另外,在土壤含水量为60%WHC时,氮添加显著降低了整个冻融期、融化期和冻结期土壤CO2累积排放量,分别降低了65.6%、64.0%和67.4%(P<0.05,图3);而在在土壤含水量为80%WHC时,氮添加对整个冻融循环期、融化期和冻结期的CO2累积排放量均无显著影响。
3 讨 论
与冻融循环前相比,19个冻融循环显著降低了土壤微生物量(图2),这与很多研究结果一致[4,35],主要是由于土壤微生物在冻融循环过程中大量死亡、土壤基质的流动性和可利用性受限,导致土壤微生物量下降[36]。有研究认为,冻结期土壤温度低于-5℃就可导致土壤微生物死亡[37]。在冻融循环条件下,土壤含水量的变化对土壤微生物量并没有显著影响,可能是由于,本研究为室内培养实验,培养前土样过筛破坏了土壤团聚结构,加之土壤样品量较少,导致冻融循环过程中冰晶的冻胀对土壤结构的破坏作用较小。在60%WHC土壤含水量时,氮添加显著增加了土壤MBC含量,说明氮添加减缓了冻融作用对土壤微生物量的降低,其原因可能是在适宜的土壤含水量条件下,氮添加促进了土壤微生物生长[15]。而在80%WHC土壤含水量条件下,氮添加显著增加了土壤MBN含量,这可能是由于在冻融条件下,水和氮素同时添加促进了氮同化微生物群落的生长,进而增加了土壤MBN含量。
在冻融循环条件下,SOC矿化释放的CO2在土壤融化期和冻结期分别占冻融循环期CO2总释放量的54%和46%,二者差异较小。首先,东北地区的土壤微生物经过长期的低温筛选,耐低温的土壤微生物可能比较丰富,导致冻结期也有一定量的土壤CO2排放。另外,本研究模拟东北黑土区实际气候条件下的冻融循环情景,冻融循环频率(24h 为1个冻融循环周期)设置较高,导致冻结期(12h)和融化期(12h)的降温和升温过程可能彼此会造成一定影响,加之土壤温度的变化一般滞后于空气温度的变化[38],最终可能掩盖了土壤有机碳矿化量在土壤融化期和冻结期的真实差异。
冻融循环可影响土壤物理性质和养分有效性的变化,因此,冻融循环过程中土壤含水量和氮素有效性等变化将影响土壤有机质的矿化过程,导致土壤CO2排放量的变化[39-40]。本研究发现土壤含水量与氮素添加对土壤有机碳的矿化有显著的交互影响(表3,图3)。在适宜的土壤含水量条件下(60%WHC),不管是冻结期还是融化期,氮添加均抑制了土壤有机碳矿化量,进而降低了整个冻融期的土壤有机碳矿化量。氮添加提升了土壤氮素有效性(图3),微生物氮矿化理论(Microbial N mining theory)认为,在氮限制的土壤中,外源无机氮素的输入会使土壤微生物对氮素的需求从土壤有机质转到外源无机氮,从而降低了微生物对土壤有机质的矿化,减少了土壤CO2排放量[41]。另外,由于氮素输入促进了微生物生长(图2),因此,在适宜的水分条件下,氮添加可能提高了微生物碳利用效率。而在较高土壤含水量条件下,氮素输入对冻结期、溶解期和整个冻融期的土壤碳矿化量都没有显著影响,这是由于,在冻融循环条件下,较高的土壤含水量可抑制土壤微生物活性,减少土壤有机碳矿化量[42-43]。在本研究中,与适宜水分条件相比,在无氮素添加时,水添加显著抑制了融化期的土壤有机碳矿化(图3),因此,氮素添加对土壤微生物的影响可能比水分的影响小,从而造成在较高土壤含水量条件下,氮素添加对培养期间土壤CO2排放的影响较为微弱。
综上,在冻融循环条件下,土壤有机碳的矿化受土壤含水量和氮素有效性的共同影响。在东北黑土区,初春和秋末土壤冻融现象明显,在该时期,黑土农田土壤水分变异较大。因此,从土壤有机碳矿化和土壤固碳的角度考虑,为降低土壤有机碳矿化,保持土壤有机碳含量,在农田管理中应综合考虑农田排水状况以及土壤氮素水平。
4 结 论
经过19个冻融循环后,黑土农田土壤有机碳的矿化量在融化期和冻结期分别占总矿化量的54%和46%。土壤含水量与氮素添加对黑土农田土壤有机碳矿化量有显著的交互影响。无论是否有氮素添加,水分添加对整个冻融期总土壤有机碳矿化量均无显著影响。在较适宜的土壤含水量条件下,氮添加显著降低了土壤有机碳矿化量,而在较高含水量条件下,氮添加对土壤有机碳矿化无显著影响。因此,在黑土农田土壤冻融期,适宜的土壤含水量和较高氮含量可减少土壤有机碳的矿化,有助于土壤有机碳的固定。
