外源草酸对绿竹笋抗氧化酶和木质化的影响
2020-06-29余学军
俞 暾,郑 剑,余学军
(1.浙江农林大学 省部共建亚热带森林培育国家重点实验室,浙江 杭州 311300;2.浙江农林大学 林业与生物技术学院,浙江 杭州 311300;3.浙江农林大学 农业与食品科学学院,浙江 杭州 311300)
绿竹Bambusa oldhami是禾本科Gramineae竹亚科Bambusoideae绿竹属Bambusa的优质笋用竹植物,分布于华南丛生竹林亚区和华中亚热带混生竹林亚区南部,5-10月为产笋期[1-3]。绿竹笋形似马蹄,俗称“马蹄笋”,笋肉细嫩,味道鲜美,深受人们喜爱。由于出笋时期温度较高,加之出笋集中,绿竹笋采后容易发生变质、腐败、木质化,储藏难度较大,限制了产品的流通和销售[4-5]。草酸广泛存在于动植物和真菌体内,具有很强的抗氧化性[6]。适当浓度的草酸溶液可以作为抗氧化剂应用于果蔬的保鲜、储藏与运输等。有研究发现:草酸处理能够有效延缓鲜切香蕉Musa nana[7]、荔枝Litchi chinensis[8]、去皮荸荠Heleocharis dulcis[9]的变质腐败,延长保鲜期。关于外源草酸处理对于采后去壳绿竹笋的抗氧化和木质化的影响尚未被报道。因此,本研究以绿竹笋为对象,研究外源草酸对去壳绿竹笋抗氧化酶和木质化的影响,为竹笋的采后储藏保鲜提供依据。
1 材料与方法
1.1 材料的选择与处理
供试材料绿竹笋于2017年7月下旬采自浙江省瑞安市,瑞安市地理坐标27°40′~28°00′N,120°10′~120°15′E,全年无严寒酷暑,冬短夏长,四季分明,雨水充沛。全年平均气温17.9 ℃,年平均降水量1 110~2 200 mm,历史年平均降水量1 527.2 mm。挑选外观完好且直径和长度相近的绿竹笋,放置于泡沫箱中6 h内运至实验室。绿竹笋样品切除基部2~3 cm不可食用部分,小心剥除笋壳,用自来水清洗干净沥干,处理组浸入5 mmol·L-1草酸溶液中10 min(该草酸浓度和处理时间为前期预实验获得的最佳处理参数),对照组浸入自来水保持同等时间,取出沥干后于阴凉通风处晾干,然后放于塑料筐中,套0.5 mm厚度的聚乙烯袋,不封口,置于(6±1) ℃的恒温恒湿箱(Sanyo,MIR-554)中储藏。处理当天取样为T0,随后每天随机取样,取样时间为Tn,n=1、2、3、4、5、6 d,取笋体基部切口位置往上2~4 cm的一段笋肉用于指标测定。每个处理每次取5根笋样,重复3次。
1.2 测定项目及测定方法
过氧化氢(H2O2)质量摩尔浓度采用苏州科铭生物技术有限公司过氧化氢试剂盒测定。超氧化物歧化酶(SOD)活性、过氧化氢酶(CAT)活性、苯丙氨酸解氨酶(PAL)活性、过氧化物酶(POD)活性参照曹建康等[10]的方法。木质素质量分数参照鞠志国等[11]的方法。4-香豆酸-辅酶A连接酶(4-CL)活性参照毕咏梅等[12]的方法并加以修改。羟基肉桂醇脱氢酶(CAD)活性参照CAI等[13]的方法并加以修改。基因表达量采用大连TaKaRa公司SYBR Premix Ex TapTM(TliRNaseH Plus)Kit测定。
1.3 统计与分析方法
用Excel进行数据统计分析及图表制作;用SPSS 22进行0.05水平差异显著性分析。
2 结果与分析
2.1 低温下草酸对采后绿竹笋过氧化氢质量摩尔浓度和抗氧化酶活性的影响
由图1A可知:草酸处理组与对照组的绿竹笋H2O2质量摩尔浓度在冷藏期间的变化趋势基本一致,总体上呈先上升后下降的趋势,草酸处理组的H2O2质量摩尔浓度在冷藏2~6 d时显著低于对照组(P<0.05),比对照组分别降低了9.2%、8.5%、14.3%、10.2%和12.2%。由图1B和C可知:草酸处理组与对照组的绿竹笋SOD和CAT活性在冷藏期间的变化趋势基本一致,总体上呈先上升后下降的趋势;草酸处理组的SOD活性在冷藏2~6 d时显著高于对照组的SOD活性(P<0.05),比对照组分别提高了23.4%、41.5%、18.1%、14.6%和16.4%;草酸处理组的CAT活性在冷藏2~6 d时显著高于对照组(P<0.05),比对照组分别提高了20.2%、7.1%、78.4%、29.7%和25.6%。由此表明:外源草酸处理可以提高绿竹笋体内抗氧化酶SOD、CAT的活性,并延缓H2O2的积累。
2.2 低温下草酸对采后绿竹笋木质素质量分数与硬度的影响
如图2可知:草酸处理组与对照组的绿竹笋木质素质量分数和硬度在冷藏期间的变化趋势基本一致,总体上呈上升趋势。由图2A可知:草酸处理组的木质素质量分数在冷藏的2、3和5 d时显著低于对照组(P<0.05),比对照组分别降低了25.3%、17.3%、12.7%。由图2B可知:草酸处理组的硬度在冷藏的3~5 d时显著低于对照组(P<0.05),比对照组分别降低了30.5%、33.8%、16.5%。由此说明:外源草酸处理可以延缓绿竹笋木质素的积累和硬度的上升。
2.3 低温下草酸对采后绿竹笋木质素合成关键酶活性的影响
如图3可知:草酸处理组与对照组的绿竹笋PAL、4-CL、CAD、POD活性在冷藏期间的变化趋势基本一致。如图3A可知:PAL活性总体上呈上升的趋势,草酸处理组的PAL活性在冷藏2~5 d时显著低于对照组(P<0.05),比对照组分别降低了23.3%、25.8%、16.0%、18.2%。如图3B可知:4-CL活性总体上呈先上升后下降的趋势,草酸处理组的4-CL活性在冷藏3 d时显著低于对照组(P<0.05),比对照组降低了26.3%。如图3C可知:CAD活性总体上呈先上升后下降的趋势,草酸处理组的CAD活性在冷藏3~5 d时显著低于对照组(P<0.05),比对照组分别降低了17.7%、17.0%、19.7%。如图3D可知:POD活性总体上呈先上升后下降的趋势,草酸处理组的POD活性在冷藏2~3 d时显著低于对照组(P<0.05),比对照组分别降低了19.9%、21.1%。由此表明:外源草酸处理可以抑制绿竹笋木质素合成关键酶PAL、4-CL、CAD、POD活性的上升。
2.4 低温下草酸对采后绿竹笋木质素合成关键酶基因表达量影响
如图4可知:草酸处理组与对照组的绿竹笋PAL、4-CL、CAD、POD基因相对表达量在冷藏期间的变化趋势基本一致。由图4A可知:PAL基因相对表达量总体上呈先下降后上升再下降的趋势,草酸处理组的PAL基因相对表达量在冷藏3~5 d时显著低于对照组(P<0.05),比对照组分别降低了37.4%、28.1%、45.4%。由图4B可知:4-CL基因相对表达量总体上呈上升的趋势,草酸处理组的4-CL基因相对表达量在冷藏3~5 d时显著低于对照组(P<0.05),比对照组分别降低了45.0%、17.5%、28.1%。由图4C可知:CAD基因相对表达量总体上呈先上升后下降的趋势,草酸处理组的CAD基因相对表达量在冷藏2~3 d时显著低于对照组(P<0.05),比对照组分别降低了34.7%、48.2%。由图4D可知:POD基因相对表达量总体上呈下降的趋势,草酸处理组的POD基因相对表达量略低于对照组,差异不显著(P>0.05)。由此说明:外源草酸处理可以抑制PAL、4-CL、CAD和POD基因的表达。
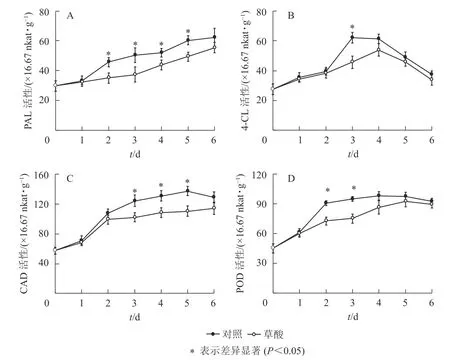
图3 低温下草酸对绿竹笋PAL、4-CL、CAD和POD活性的影响Figure 3 Effects of oxalic acid treatment on PAL, 4-CL, CAD and POD activities of bamboo shoots at low temperature

图4 低温下草酸对绿竹笋PAL、4-CL、CAD和POD基因相对表达量的影响Figure 4 Effects of oxalic acid treatment on PAL, 4-CL, CAD and POD relative expression of bamboo shoots at low temperature
3 讨论与结论
本研究表明:外源草酸处理可以提高绿竹笋体内抗氧化酶SOD、CAT的活性并延缓H2O2质量摩尔浓度的积累;可以延缓木质素的积累和硬度的上升;既可以抑制绿竹笋木质素合成关键酶PAL、4-CL、CAD、POD活性的上升,又可以抑制PAL、4-CL、CAD和POD基因的表达;可以减缓绿竹笋的木质化进程,延缓绿竹笋品质的下降,延长绿竹笋的保鲜期。
植物在受到外界逆境胁迫时会产生活性氧(ROS),导致细胞膜脂过氧化,破坏生物膜的结构和功能,严重则会致使细胞程序性死亡。为了抵御ROS的伤害,植物体会激活体内抗氧化防御系统,提高抗氧化酶的活性,以消除ROS[14-17]。本研究结果显示:在低温储藏下,草酸处理组的去壳绿竹笋笋肉组织中的H2O2质量摩尔浓度显著低于对照组(P<0.05),较低的H2O2质量摩尔浓度有助于减轻ROS对笋肉细胞的毒害。草酸处理组的去壳绿竹笋笋肉组织中的SOD和CAT活性显著高于对照组(P<0.05)。梁春强等[18]研究认为:适合浓度的草酸处理不但可以显著提高氧化酶SOD和CAT的活性,还可以有效降低O2-的生成速率和H2O2的含量。
木质素为植物次生代谢的产物,属酚类化合物,是构成细胞壁次生结构的主要成分。竹笋在采后非常容易老化变质,失去食用价值,其中一个重要的因素就是木质化。木质化主要表现为笋体组织硬度上升,木质素质量分数增加,笋体发生褐变等[19-21]。除了竹笋,茭白Zizania latifolia、芦笋Asparagus officinalis等蔬菜在采后储藏过程中均有木质化的发生[22-25]。本研究发现:低温条件下储藏,草酸处理不但可以抑制绿竹笋硬度的上升,还能够减缓木质素的生物合成,抑制采后去壳竹笋的木质化,维持较好的口感品质,延长竹笋可食用期限。与王琪等[26]研究的采用外源草酸抑制带壳雷竹Phyllostachys praecox笋木质素的合成累积以及硬度的上升的结论相似。
PAL、4-CL、CAD和POD等4种酶与木质素的合成有密切的关系,其活性的升高都能促进木质素的合成,提高组织的木质化程度。本研究结果表明:低温条件下,经草酸处理的绿竹笋的PAL、4-CL、CAD和POD的活性均低于对照组,草酸处理在总体上明显抑制了去壳绿竹笋的木质素生物合成关键酶的活性。这与沈玫等[27]研究的利用外源草酸抑制带壳绿竹笋PAL和POD活性的结论类似。
果蔬的木质化进程受编码木质素代谢的关键酶基因的表达调控。LU等[28]研究发现:PpCAD1和PpCAD2这2个基因主要调控梨Pyrus pyrifolia的木质素代谢生物合成。不同种类的果蔬,调控木质素合成关键酶的基因也不同,SHAN等[29]研究表明,调控枇杷Eriobotrya japonica的木质素代谢基因主要是EjCAD1、EjPOD、EjPAL2这3个基因。此外,大多数研究人员认为是PAL、4-CL、CAD和POD的活性与其对应基因的表达在协同调控木质素的合成[30-31]。本研究结果表明:草酸处理在冷藏的中后期能够显著抑制去壳绿竹笋的木质素合成关键酶PAL、CAD和4-CL的基因表达水平,可以抑制POD的基因表达水平。结合硬度、木质素质量分数以及对应酶的活性的变化趋势分析可见,PAL、4-CL、CAD基因表达与硬度、木质素质量分数以及酶活性的变化趋势相一致,POD基因表达与硬度、木质素质量分数以及酶活性的变化趋势相反,说明绿竹笋木质素合成关键酶的基因表达和活性变化同步调控木质素的生物合成。LU等[28]研究发现:利用氯化钙可以抑制梨果肉中PpCAD1和PpCAD2基因表达,从而调控木质素的生物合成途径,最终达到抑制梨的硬化的效果,这与本研究结果类似。另外,SEWALT等[32]研究发现,烟草Nicotiana tabacum的PAL基因在受到抑制之后,不但木质素质量分数明显下降,PAL的活性也明显降低。本研究中草酸处理既抑制了木质素代谢关键酶活性,又抑制了对应基因表达水平,从2个方面抑制了去壳绿竹笋笋肉硬度的上升和木质素的累积,延缓了冷藏过程中竹笋食用品质的下降。
