山西灵空山油松径向生长对气候变化的响应*
2020-06-28许玲玲同小娟张劲松
许玲玲,同小娟**,张劲松,孟 平,王 鑫 ,李 俊
(1.北京林业大学林学院,北京 100083;2.中国林业科学研究院林业研究所国家林业局林木培育重点实验室,北京 100091;3.中国科学院地理科学与资源研究所陆地水循环及地表过程重点实验室,北京 100101)
随着工业的发展、森林的过渡采伐、化石燃料的燃烧和土地利用方式的改变,大气CO2浓度呈不断升高趋势[1]。长期以来,气候变化对森林生产力的影响始终是生态学和林学的研究热点之一,尤其在全球变暖的大背景下,研究过去长时间序列里气候因子对生产力的影响机制,对了解并预测未来气候条件下的森林生长具有重要意义[2]。最新数据显示,近5a 中国新造人工林4.47 亿hm2,比5a 前增长了21.31%,中国人工林总面积已位居全球之首[3],但人工林面临着质量不高、结构不合理、立地条件差等问题[4],在此背景下研究过去气候对人工林生长的影响,可为预测未来气候变化条件下人工林生长变化提供重要依据。
油松(Pinus tabuliformis)分布于吉林南部、辽宁、河北、河南、山东、山西、内蒙古、陕西、甘肃、宁夏、青海及四川等省区,生于海拔100-2600m地带,其具有适应性强,根系发达,耐干旱瘠薄等特点,可以作为荒山造林的先锋树种,在北方干旱地区造林中得到广泛应用。树木年轮以其定年准确、连续性好、分辨率高、样本易获得以及对环境变化敏感性强等优势,已成为对全球变化特别是对过去全球气候与环境变化研究的重要基础资料[5-6]。因此,利用树木年轮分析方法研究气候变化对森林的影响具有极其重要的生态学和实际意义[7]。
国际上常用树木年轮学研究气候变化和树木生长的关系以及重建气候序列[8-11]。Metsarant 等[12]利用加拿大温哥华冷杉林(Abies fabri)的年轮数据,分析了森林的生长动态,发现模型计算出来的生产力与实际计算的生产力存在差异。Vieira 等[13]研究了不同生长阶段海岸松(Pinus pinaster)年轮以及密度对地中海气候的响应,发现幼龄期早材宽度对气候响应更敏感,而老龄生长阶段晚材宽度对气候的响应更强。Bogino等[14]分析了海岸松和欧洲赤松(Pinus sylvestris)1975-1999年树轮稳定碳同位素与水分利用率,表明海岸松的水分利用率与冬季和春季的相对湿度呈负相关关系。张朋磊等[8]建立了不同纬度兴安落叶松(Larix gmelinii)年轮宽度年表,并分析了不同纬度气候因子变化规律以及气候因子对年表的影响,发现随着纬度的升高,气温升高对落叶松的径向生长所产生的作用由抑制转为促进。油松由于其年轮变化线清晰明显,可进行可靠的年轮分析和定年,是用于气候与环境重建的重要树木年代学代用资料,所以被许多学者作为研究对象探讨气候与树木生长的关系[15-18],然而针对不同生长阶段林分对气候响应的相关研究并不多,对于林分生长阶段、林木大小、林分密度以及其它因素引起的树木径向生长尚需深入研究。
本研究以树木年轮学为基础,以灵空山不同生长阶段油松树为研究对象,建立了灵空山不同生长阶段油松树轮标准化年表,通过对影响油松生长的气候因子进行Pearson 相关分析,获得各气候因子与油松年轮宽度的相关性及响应函数,明确对油松年轮宽度生长有重要影响的气候因子,以期为深入研究该区域气候变化及灵空山油松对气候变化的响应提供科学参考。
1 材料与方法
1.1 研究区概况
试验地点山西灵空山国家级自然保护区灵空山林场(36°67′44″N,112°09′80″E),位于山西沁源县西南部及古县北部,地处太岳山脉中段的深山腹地,属温带大陆性气候,四季分明,年平均气温6.2℃,日照时数2600h,降水量662mm,无霜期145d 左右,海拔1583-1660m(平均1618m)。该地区植被为以油松为主的温性针阔混交林,群落乔木优势种有油松、辽东栎(Quercus wutaishanica),伴生树种有白蜡(Fraxinus chinensis)、杜梨(Pyrus betulifolia)、漆树(Toxicodendron vern icifluum)等阔叶树;灌木有黄刺玫(Rosa xa nthina)、土庄绣线菊(Spiraea pubescens)、金花忍冬(Lonicera chrysantha)等;草本植物主要有披针苔草(Carex lanceolata)、披碱草(Elymus dahuricus)、苍术(Atractylodes lancea)等。
1.2 样品的采集与处理
2019年6月,按照国际年轮数据库(ITRDB)的标准,在海拔1503-1524m 油松生长良好且未间伐区域内,设置20m×20m 的油松人工林样地,样地为西南坡,凸形坡,坡度37°~43°,郁闭度0.72,每木检尺结果为油松平均胸径24.32cm,密度为1125棵·hm-2。在样地内选取20 棵健康优势树,利用生长锥在胸径(1.3m)高处向东西和南北两个方向垂直钻取两根树芯,树芯采集后将样品及时放入吸管中保存并编号记录好样点信息,采集结束后,使用树木生长愈合剂对其进行涂抹,防止虫蚁蛀食。共获得20 棵树的40 根树芯,基本可以反映样地的油松生长情况。样本采集后带回实验室,放置于通风处阴干,将样本固定于木条上并使用200、400 和600目砂纸依次打磨,直至油松年轮在肉眼下清晰可辨。
1.3 制作年表
1.3.1 年表制作方法
使用Lintab6 结合TASP-Win 软件测量年轮宽度,精确至0.01mm,在双筒显微镜下用骨架法进行目视交叉定年,使用COFECHA 软件对初步测量的树轮宽度进行交叉定年检验,根据COFECHA 交叉定年程序检验测量结果,可以对测量错误或定年错误进行修改,把主序列相关性差的样芯剔除,确保定年和测量准确,检验出来定年错误的样芯用年轮分析系统重新测量树轮宽度,并用COFECHA 软件再次进行交叉定年。用ARSTAN 软件对相关性高的序列进行年轮宽度的标准化以及年轮宽度年表制作,由于树木的生长不仅受环境和生态因子的影响,而且还受到自身生理因子的控制,所以通过消除非气候因素和树木生长趋势的影响即树轮去趋势处理,进而得到标准化年表(standard chronology,STD)。ARSTAN 软件去趋势的原理是利用一定的曲线模型来拟合树木生长趋势,利用年轮序列与拟合曲线序列的比值得到去趋势序列。年表中的年轮宽度指数(RWI)为实际测得年轮宽度与去趋势后的年轮宽度值的比值。
参照林业上对油松人工林生长阶段级和龄期的划分标准,可知0~20a为幼龄期(young-aged forest);21~30a 为中龄期(middle-aged forest),31~40a 为近熟期(near mature forest);41~60a 为成熟期(mature forest);61a 以上为过熟期(over mature forest)。
树木年代学研究中,有一些常用的年表统计参量,包括年轮宽度指数、信噪比、平均敏感度、样本间相关系数、样本总体代表性、样本一阶自相关系数。
1.3.2 年表统计参量
(1)年轮宽度指数
在树木年轮宽度年表建立的过程,采用统计学方法对年轮宽度序列进行曲线拟合(样条函数,双曲线,多项式,指数函数),得到树木生长的期望值(Yi),年轮宽度指数(Ii)是树木年轮实际宽度值(Wi)与期望值(Yi)之比。

(2)信噪比
信噪比(SNR,signal-to-noise ratio)是树轮年表中气候信息与非气候噪声的比值,信噪比数值越大,年表中所包含的气候信息越多,可用来衡量样本中包含的环境信息量的大小,表达式为

(3)样本一阶自相关系数
样本一阶自相关系数(AC,autocorrelation)表示上一年的气候因子对当年年轮生长的影响,一阶自相关系数越大,说明上一年的气候因子对当年年轮生长的影响越大,反之,则表明影响较小。
(4)样本间相关系数
样本间相关系数(mean inter-series correlation,rbar)是不同样本序列间年轮宽度变化一致性大小的统计量。样本间相关系数越大,样本序列间相似度越高,表明样本序列受到相似的环境气候因子影响,样本序列间年轮宽度变化较一致。
(5)平均敏感度
平均敏感度(MS,mean sensitivity)是衡量年轮宽度逐年变化的统计量,平均敏感度越大,树木生长受气候因子的限制作用就越大,大部分研究中,质量较好的年表的平均敏感度取值范围在0.1~0.6,大部分数值在0.1~0.4,表达式为
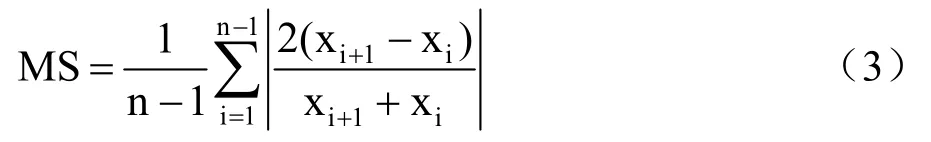
式中,xi为第i 个年轮宽度值;xi+1为第i +1 个年轮宽度值;n 为样本年轮总数。
(6)第一主成分所占方差量
第一主成分所占方差量(variance in first eigenvector)是反映年表中各样本序列同步性大小的统计参数,其值越大,气候因子对树木年轮宽度生长的限制作用越大。
1.4 气候数据
气候数据来源于CRU TS 4.02 全球气候数据库,其网格点的间隔为0.5°×0.5°,选择了距离采样点经纬度最近且海拔差距最小的网格点的气候数据作为本研究使用的气候数据。经过整理后得到灵空山林场1951-2018年的降水量、平均气温、最高气温、最低气温、干旱指数、以及气温幅值的逐月数据和逐年数据。干旱指数为月尺度的帕默尔干旱指数(PDSI),气温幅值为月尺度平均最高气温与平均最低气温的差值。研究区属于温带大陆性气候,雨热同期现象明显,夏季高温多雨,冬季寒冷干燥,从降水的季节分布来看,一年中的大多数雨量集中在夏季且气温与降水量的变化趋势类似。考虑到气候因子的“滞后效应”,选择上年9月-当年10月的气候数据进行分析。
1.5 数据分析
采用Mann-Kendall(M-K)趋势检测法检验1951-2018年灵空山不同生长阶段各气候因子的变化[19],同时对不同生长阶段各气候因子的年际变化进行线性拟合。采用Pearson 相关分析法研究不同生长阶段树木年轮宽度指数与逐月月平均气温、降水量、最低气温、最高气温、气温幅值、干旱指数的相关关系,分析不同月份气候对油松生长的影响及不同时期的主导因素。
利用Origin2018 作图,利用SPSS 分析统计年表和气候的相关性。
2 结果与分析
2.1 树轮宽度年表统计特征分析
表1 为灵空山油松标准化年表的各项基本统计特征。由表可以看出,5 个龄期油松序列与主序列的平均相关系数分别为0.447、0.475、0.349、0.447、0.515,均在0.4 左右波动,表明取样点的样本序列与主序列的同步性较好。不同生长阶段油松样本序列的一阶自相关系数均达到0.5 以上,其中成熟期达到0.734,说明油松的径向生长受到上一年气候因子的影响作用较大,而这一现象在成熟期表现最为明显。平均敏感度用来度量相邻年轮之间年轮宽度的变化情况,5 个龄期的平均敏感度分别为0.338、0.324、0.308、0.297、0.354,从不同龄期油松年表特征可以看出,平均敏感度、信噪比等指标均较高,说明所建立的年表包含较多的气候信息,可以用于年表与主要气候因子的相关性分析。各龄期的油松样本总体代表性均达到了0.85 的可接受水平,分别为0.931、0.934、0.938、0.961、0.96,且第一主成分所占方差量均较大,说明适用于气候因子的响应分析。从灵空山油松年轮宽度标准化年表(图1)可以看出,树轮宽度随着时间序列发生变化且存在频繁波动。
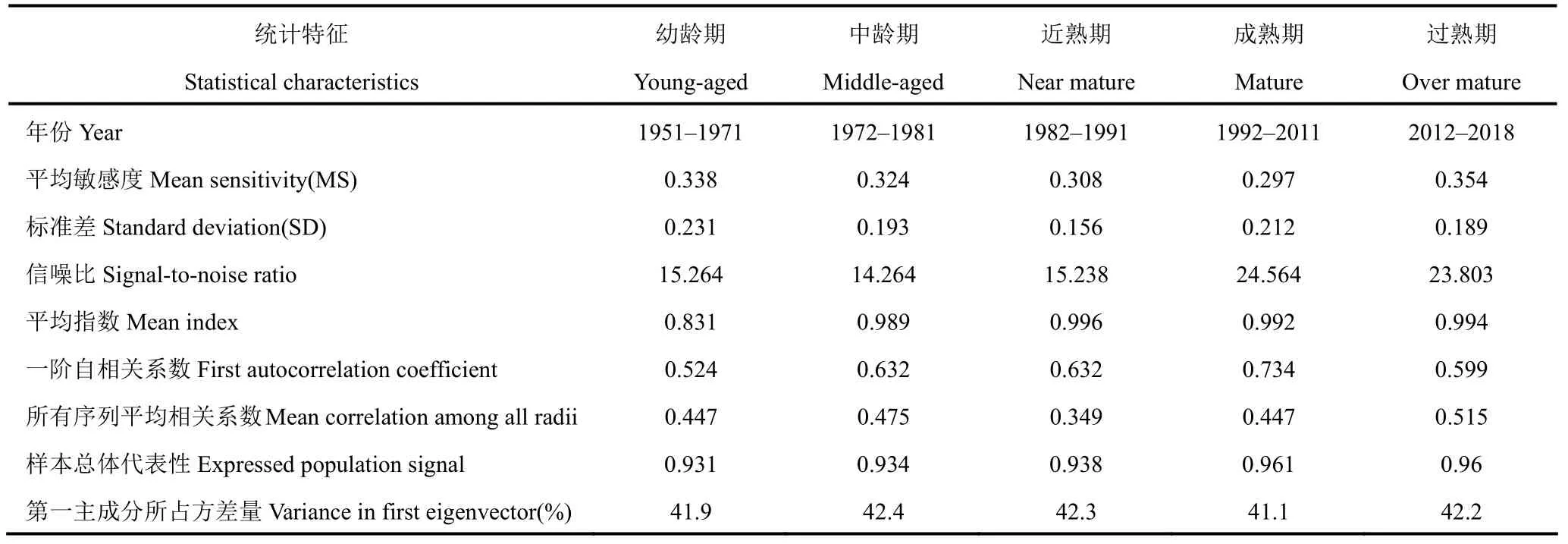
表1 灵空山油松树轮宽度年表的统计特征Table 1 Chronology statistics of the ring-width of P.tabulaeformisin the Lingkong Mountain
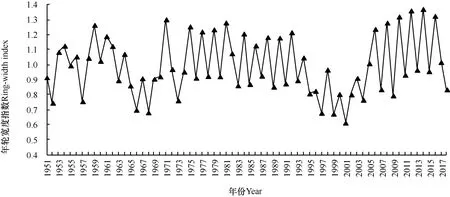
图1 灵空山油松年轮宽度标准化年表(1951-2018年)Fig.1 Standard chronology of ring width of Pinus tabulaeformis in the Lingkong mountain(1951-2018年)
2.2 同期气候因子变化特征分析
图2 为1951-2018年年平均降水量、平均气温、最高气温、最低气温、气温幅值以及干旱指数的变化。统计分析发现,1951-1971年油松幼龄期间,研究区年平均气温、平均最高气温、气温幅值以及干旱指数均有下降,但变化趋势不显著,其中平均气温以及平均最高气温分别以0.013℃·a-1、0.033℃·a-1的速率下降,降水量以及最低气温总体有上升,但趋势不显著。在1972-1981年油松中龄期间,平均气温的上升速率为0.022℃·a-1,降水量的下降幅度为0.109mm·a-1,但变化趋势均不显著。1982-1991年油松近熟期间,除降水量以0.299mm·a-1的速率下降外,其它气候因子均有所上升,但亦均未达显著水平。1992-2011年油松成熟期间,年降水量、平均气温以及最高气温变化均不显著,但最低气温以及气温幅值的变化均达到极显著水平(P<0.01),最低气温以0.06℃·a-1的速率上升,气温幅值的下降速率为0.05℃·a-1。2012-2018年油松过熟期间年降水量、平均气温、最高气温、最低气温以及干旱指数均呈上升趋势,其中平均气温以及最低气温的上升幅度分别为0.135℃·a-1、0.17℃·a-1,均达到了极显著水平(P<0.01)。
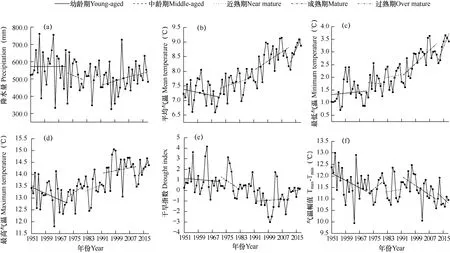
图2 1951–2018年年降水量、平均气温、平均最高气温、平均最低气温、气温幅值以及干旱指数的变化Fig.2 Changes of annual precipitation, average temperature, average maximum temperature, average minimum temperature, -and drought index in 1951-2018
2.3 气候变化对油松年轮宽度的影响
由于油松年轮生长可能受上年气象因子影响[20-22],因此,选择各阶段上一年9月-当年10月的月平均气温、月平均最高气温、月平均最低气温、月降水量、月气温变幅以及月干旱指数与油松年轮宽度标准化年表分别进行相关性分析,结果见图3。由图可见,油松幼龄期径向生长与当年8月降水量呈极显著正相关关系(P<0.01),与6月最高气温和气温幅值均呈显著负相关关系(P<0.05)。近熟期的径向生长与上年10月降水量呈显著正相关,而与当年10月降水量呈显著负相关关系,且油松的径向在生长后期与降水的相关系数均大于幼龄期与中龄期。成熟期径向生长与7月和8月的降水量呈显著正相关。过熟期的径向生长与上年11月降水量呈显著正相关,而与当年1月和5月的降水量分别呈极显著和显著负相关,且相关系数均达到0.7 以上。
由图3 可见,油松的径向生长在不同的生长阶段对气温的响应程度也各不相同。幼龄期的径向生长与当年6月最高气温呈显著负相关,与当年2月和10月最低气温分别呈极显著和显著正相关,与6月气温幅值呈显著负相关。中龄期的径向生长与7月平均最低气温呈极显著正相关关系,与1月气温幅值呈显著正相关而与7月气温幅值呈极显著负相关。近熟期的径向生长与4月最低气温和上年10月气温幅值呈显著负相关,与上年12月气温幅值呈显著正相关。成熟期的径向生长与当年1月平均气温呈显著负相关,与1月最高气温呈极显著负相关,与3月最高气温呈显著负相关,与上年12月和1月气温幅值呈显著负相关。过熟期的径向生长与2月和8月最高气温呈显著负相关,且相关系数达到0.7,明显高于其它龄期,与月平均气温、最低气温以及气温变幅的相关关系均未达到显著水平。
幼龄期径向生长与8月和10月干旱指数呈显著正相关,近熟期径向生长与7月和8月干旱指数分别达到了极显著与显著正相关水平;过熟期径向生长与上年11月以及当年4月干旱指数显著正相关。可见,树轮宽度受外界环境因素的影响随时间序列产生不同的变化趋势。
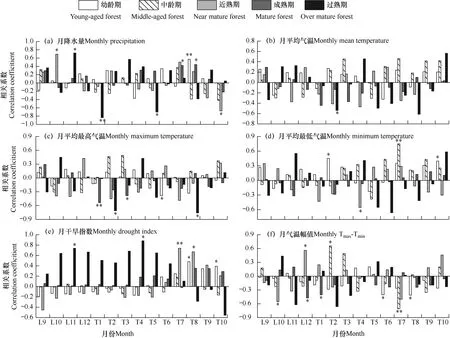
图3 不同生长阶段油松标准化年表与月平均气候因子之间的相关关系(1951-2018)Fig.3 Correlation between standardized chronology of Pinus tabulaeformis and monthly climate factors in different growth stages (1951-2018)
3 讨论与结论
3.1 讨论
油松5 个龄期的平均敏感度均在0.3 左右,信噪比、标准差较高以及一阶自相关系数均较高,表明树木年轮年表中含有较高的气候信息且油松年轮宽度的生长受上一年气候因子的影响较大,油松年轮宽度的生长具有滞后效应[23],样本总体代表性达到了0.85 可接受水平,适用于进行年轮气候学分析,由标准化年表可知,各龄期第一主成分所占方差量均较大,适用于气候因子的响应分析[24],过熟期与成熟期的信噪比高于其它龄期且数值大小较一致,说明过熟期与成熟期包含有更多的气候信息,且在这两个龄期内油松的径向生长无明显差异[25],不同生长阶段油松的径向生长对气候响应敏感度不同,其中幼龄期与中龄期的敏感性大于近熟期和成熟期,这与Wu 等[26]研究的不同生长阶段天山云杉的径向生长对气候的响应结果一致,但是也有研究发现树木随着生长阶段的增长,水分运输速率会下降,使过熟期径向生长对气候的敏感性更大[27-28],导致有关生长阶段对树木生长与气候关系的研究结果和结论在一些树种和地区上出现差异。
不同生长阶段油松的径向生长对气候的响应程度不同。Wang 等[29]对中国东北不同生长阶段兴安落叶松的径向生长作了相关研究,研究结果表明落叶松的径向生长对气候的响应随生长阶段的变化而显著变化。Yu 等[30]对祁连山不同生长阶段的刺柏进行了研究,发现老龄刺柏的径向生长比幼龄刺柏对气候响应更为敏感,相似的研究结论也被发现于地中海气候的巴西栎中[31]。研究表明,中国东部地区油松径向生长主要受到夏季温度和降水的影响[8]。从生理学角度讲,春季树木开始生长,降水增加,则早材细胞分裂快,细胞数量多且体积大,所以易形成宽年轮[32]。温度和降水对树木的影响是两者综合影响的结果,降水增加,阴雨天时间加长,光照不足,温度下降,树木光合作用速率下降,细胞分化推迟,纤维减少,细胞壁变薄[33]。
由油松幼龄期径向生长与月气候因子的相关性分析可以看出,年轮宽度与降水量多呈正相关关系,其中与当年8月降水量呈显著正相关。王亚军等[32]采集了宁夏罗山上限和下限的油松树轮样本,分别建立了年表,发现该地区的油松年表与当年3、5月的降水量呈显著正相关。崔明星等[34]对木兰围场地区、赤峰围场地区的树木与气候因子的相关性研究均发现,树木生长与气温具有显著负相关关系,与降水具有正相关关系。
近熟期的径向生长与上年10月降水量呈显著正相关,而与当年10月降水量呈显著负相关。这是由于上年冬季长期气团所控制的持续影响,当地空气湿度小、土壤水分含量低,天空晴朗少云,太阳辐射增强,气候干燥多风,土壤中的水分对树木生长起着极为关键的作用。此时,降水的增加有利于树木的径向生长,上年的充足降水也可以降低干旱胁迫的危险,并有助于增强光合速率,促进树体内糖分的合成和贮存,为下一年生长积累能量[35]。张芬等[23]研究祁连山东部不同生长阶段油松径向生长对气候的响应也得出相同结论,不同生长阶段组油松树轮指数与当年9、10月温度呈正相关,与当年8-10月的降水量呈负相关关系,过熟期相关性达到了显著水平。
成熟期油松年轮宽度与上年生长后期的平均气温、最高气温以及最低气温均呈明显的负相关关系。这可能是由于冬季树木的光合作用已经停止,冬季温度高时树木的呼吸作用和代谢作用较高,加速了树木体内供翌年生长所储存的营养物质的消耗,进而对树木生长产生不利的影响[8]。过熟期径向生长与当年1月和5月降水量呈显著负相关。1月是油松生长的萌芽期,此时油松树木生长在外形上虽不明显,但是树木体内的生命活动仍在进行。缺水时,树木叶片萎蔫,气孔关闭,二氧化碳扩散阻抗增加,光合作用受到明显抑制,植物根系活动相对减弱,树干本身细胞的分裂、伸长都受到抑制[33],成熟期油松年轮宽度与5月降水量呈显著负相关。这主要是由于油松已经基本完成了树木形成层细胞的分裂和生长,开始进入光合作用产物积累阶段,降水较多,导致云层较厚,阻止太阳光线,减缓光合作用,从而不利于油松生长[36]。成熟期油松年轮宽度与当年2月和8月最高气温呈显著负相关。在油松生长季温度的升高会导致土壤水分的蒸发量增大,影响降水对土壤水分的补给,间接表现为气温与年轮宽度呈负相关[37]。
在油松幼龄期和中龄期时,年轮宽度标准化年表与温度的相关系数均高于与降水量的值,而在成熟期则相反。这说明温度是影响幼龄期和中龄期径向生长的主要因子,而降水则是近熟期、成熟期以及过熟期木质部生长的主要制约因子。Sanchez-Salguero 等[38]对亚欧大陆不同生长阶段钩松林线对温度敏感性响应的变化发现,生长阶段对温度的响应也取决于气温变化的速率,但整体上幼龄期比老龄生长阶段对温度的敏感性更强。在本研究中,研究区油松人工林样地还存在坡度、林分密度等的差异,其中密度对于人工林胸径和材积的生长过程影响也十分明显,这些因子对油松对气候因子的响应过程有何影响以及最稳定的人工林密度等问题,还需进一步深入研究。
3.2 结论
(1)不同生长阶段油松的径向生长对气候响应敏感性不同,过熟期敏感性最大,而幼龄期和中龄期的敏感性大于近熟期和成熟期。信噪比、所有序列平均相关系数、平均指数等参数中,过熟期均大于其它龄期,过熟期更适合用于年轮气候学研究。
(2)幼龄期油松的径向生长与当年6月最高气温以及气温幅值呈显著负相关,与当年2月和10月的最低气温分别呈极显著和显著正相关,与8月降水量呈极显著正相关关系。中龄期油松的径向生长与7月平均最低气温呈极显著正相关关系,与1月的气温变幅呈显著正相关。近熟期油松的径向生长与上年10月降水量呈显著正相关,而与当年10月降水量呈显著负相关关系,且油松的径向在生长后期与降水的相关系数均大于幼龄林和中龄林。成熟期油松径向生长与7月和8月的降水量呈显著正相关;过熟期的径向生长与上年11月降水量呈显著正相关,与当年1月和5月的降水量分别呈极显著和显著负相关关系,且相关系数均达到0.7 以上。
(3)在油松幼龄期和中龄期,年轮宽度标准化年表与温度的相关系数均高于与降水量的值,而在成熟期则相反。说明温度是影响幼龄期和中龄期径向生长的主要因子,而降水量则是近熟期、成熟期以及过熟期木质部生长的主要制约因子。
