巴氏新小绥螨对芒果茶黄蓟马的功能反应研究
2020-06-19周炀陈俊谕蔡笃程符悦冠
周炀 陈俊谕 蔡笃程 符悦冠
摘 要:为明确巴氏新小绥螨(Neoseiulus barkeri Hughes)本研究对芒果上茶黄蓟马(Scirtothrips dorsalis Hood)的控害潜能,在室内温度(27±1)℃、RH (75±5)%的条件下,研究了不同虫态巴氏新小绥螨对茶黄蓟马的捕食效能,并通过其瞬时攻击率(a)、处理时间(Th)和捕食效能(a/Th)等参数进行综合评价。结果表明:巴氏新小绥螨对茶黄蓟马的成虫、1龄和2龄若虫均可取食,并具有一定的捕食能力。随着猎物密度的增加,捕食螨日取食量逐渐增加,其中巴氏新小绥螨雌成螨和第2若螨对茶黄蓟马1龄若虫的捕食效能最大,每日分别达10.35、12.82头,而雄成螨对茶黄蓟马2龄若虫的捕食效能最大,为每日12.48头。巴氏新小绥螨的雌成螨和雄成螨均对茶黄蓟马的2龄若虫表现出最强的瞬时攻击率,分别为1.0227和1.1208,第2若螨则对茶黄蓟马的雌成虫表现出最大攻击率,为1.5774。
关键词:巴氏新小绥螨;芒果;茶黄蓟马;功能反应;捕食效能
中图分类号:S436.67 文献标识码:A
Functional Response of Neoseiulus barkeri to Scirtothrips dorsalis Hood of Mango
Zhou Yang1, Chen Junyu2*, Cai Ducheng1*, Fu Yueguan2
1. College of Plant Protection, Hainan University, Haikou, Hainan 570228, China; 2. Institute of Environment and Plant Protection, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan 571101, China
Abstract: In order to determine the damage control potential of Neoseiulus barkeri on Scirtothrips dorsalis Hood of mango, the predation ability of N. barkeri on S. dorsalis Hood of mango was observed under the conditions of temperature of (27±1)℃ and RH (75±5)% in the room. The predation efficiency of different insect states of N. barkeri on S. dorsalis Hood was comprehensively assessed by the instantaneous attack rate (a), handing time (Th), predationcapacity (a/Th) and other parameters. N. barkeri could feed on the adults, nymphs of the first and second age of S. dorsalis Hood, and had a certain predating ability. With the increase of prey density, the daily feeding amount of predatory mites increased gradually. Among them, the female adult mite and deutonymph mite had the largest predationcapacity to the nymph of the first age of S. dorsalis Hood (10.35 and 12.82 per day, respectively), while the male adult mite had the largest predationcapacity to the nymph of the second age of S. dorsalis Hood (12.48 per day). The female and male adult mites of Apterygium barbati showed the strongest instantaneous attack rate to the 2-year nymph of the S. dorsalis Hood (1.0227 and 1.1208, respectively), while the deutonymph mite showed the maximum instantaneous attack rate to the female adults of S. dorsalis Hood (1.5774).
Keywords: Neoseiulus barkeri; mango; Scirtothrips dorsalis Hood; functional response; predatory efficiency
DOI: 10.3969/j.issn.1000-2561.2020.05.021
茶黃蓟马(Scirtothrips dorsalis Hood)属缨翅目(Thysanoptera)蓟马科(Thripidae)。在我国,茶黄蓟马作为芒果上的主要害虫分布于各个种植区,如海南、广西、广东、云南、福建[1]。茶黄蓟马可为害芒果嫩稍、花穗和小果,其危害可导致嫩叶失绿脱落,树体长势下降甚至停止生长,为害果实导致芒果落果及品质下降等[2]。目前对于芒果蓟马的防治手段,包括农业防治、理化诱控和化学防治等,如劳有德等[3]报道可通过农事操作控制茶黄蓟马的发生;陈永森等[4]通过悬挂蓝色诱虫板结合诱剂,发现对茶黄蓟马具有较好的诱杀效果。但是目前主要仍是以传统的化学防治方法为主,化学防治普遍存在用药频率高、剂量大等突出问题,尤其是海南地区。生物防治可实现生态环保无污染且达到可持续控制害虫的效果,但是目前在我国利用天敌进行芒果蓟马的生物防治尚未见报道。
巴氏新小绥螨(Neoseiulus barkeri)隶属于植绥螨科(Phytoseiidae)钝绥螨属(Amblyseius)。该捕食螨广泛分布于世界各地,我国北京、广州、福建、云南、江西、湖南、河北等大部分省市均有分布[5]。巴氏新小绥螨是多食性捕食者,捕食范围广,除了捕食叶螨和蓟马外,还捕食蚜虫、木虱、粉虱、介壳虫、跳虫、丝状菌、线虫和蚊蝇类幼虫等生物,植物花粉和昆虫蜜露也是其补充食物[6-8]。巴氏新小绥螨捕食量相对比较大,且容易实现人工大量扩繁,因此被广泛应用于害虫生物防治中[9]。在澳大利亚、欧洲等国家,巴氏新小绥螨已被广泛应用于防治叶螨和蓟马[10],但尚未见利用巴氏新小绥螨对茶黄蓟马进行防治的报道。本研究旨在明确利用巴氏新小绥螨对茶黄蓟马进行防控的可行性,为进一步用于茶黄蓟马的绿色防控奠定基础。
1 材料与方法
1.1 供试虫源
茶黄蓟马采自海南省乐东县佛罗镇的芒果园,该果园种植芒果品种为‘台农1号,将采集的蓟马带回室内,在養虫笼中以芒果叶片进行饲养。
巴氏新小绥螨由中国热带农业科学院环境与植物保护研究所自主扩繁。
1.2 方法
试验采取2 cm×2 cm的PVC平板,PVC平板中间开一个直径2 cm、深度0.5 cm的圆孔,平板上下均采用2 cm×2 cm、厚度0.1 cm的PVC平板覆盖,防止巴氏新小绥螨和茶黄蓟马逃跑。挑取1头巴氏新小绥螨雌成螨进入以上装置中进行24 h饥饿处理后,分别挑取以下密度的茶黄蓟马分别进入装置中:1龄若虫、2龄若虫和雌成虫2、4、6、8、10头,雄成虫3、4、5、6、7头。挑取1头巴氏新小绥螨雄成螨进入以上装置中进行24 h饥饿处理后,分别挑取以下密度的茶黄蓟马分别进入装置中:1龄若虫、2龄若虫和雄成虫2、4、6、8、10头,雌成虫1、2、3、4、5头。挑取1头巴氏新小绥螨第2若螨进入以上装置进行24 h饥饿处理后,分别挑取以下密度的茶黄蓟马分别进入装置中:1龄若虫和2龄若虫2、4、6、8、10头,雌成虫和雄成虫1、2、3、4、5头。24 h后观察并记录茶黄蓟马被捕食量的情况。以上装置试验全程置于温度(27±1)℃,湿度(75± 5)%,光周期L∶D=12∶12的人工气候箱中,每个处理重复10次。
Holling-Ⅱ型功能反应模型[11]:
Na=aTN/(1+aThN)
式中,Na为猎物被捕食数量;a为瞬时攻击率;T为试验总时间(本研究中T为1 d);N为猎物的初始密度;Th为处理时间,最大日捕食量为1/Th,捕食效能为a/Th。
寻找效应:S=a/(1+aThN)
式中,S为寻找效应;N为猎物的初始密度;a为瞬时攻击率;Th为处置1头猎物所需的时间。
1.3 数据处理
采用差异显著性分析Excel 2010和SPSS 17.0软件进行数据统计和分析,采用Duncan新复极差法进行差异性分析。
2 结果与分析
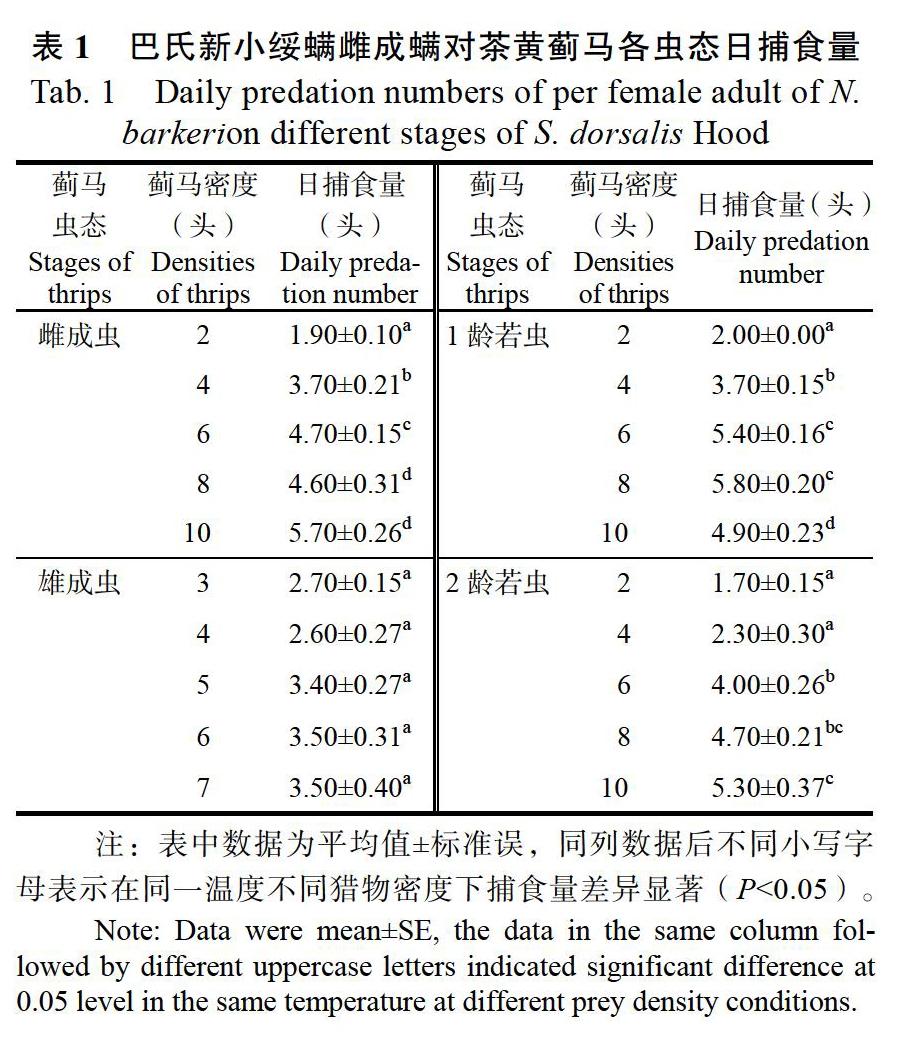
2.1 不同虫态的巴氏新小绥螨对茶黄蓟马的日均捕食量
巴氏新小绥螨雌成螨对茶黄蓟马各虫态的捕食量存在一定的差异。由表1可知,同一虫态下巴氏新小绥螨雌成螨对茶黄蓟马的日捕食量总体趋势为随蓟马密度的增加而增加,但是对于蓟马雄成虫,猎物密度的变化对于捕食螨的取食量影响差异不显著。对1龄蓟马在密度达到8头时日捕食量最大,其日捕食量与蓟马在2、4、10头密度下的日捕食量差异显著(P<0.05),与6头密度时的日捕食量差异不显著(P>0.05)。
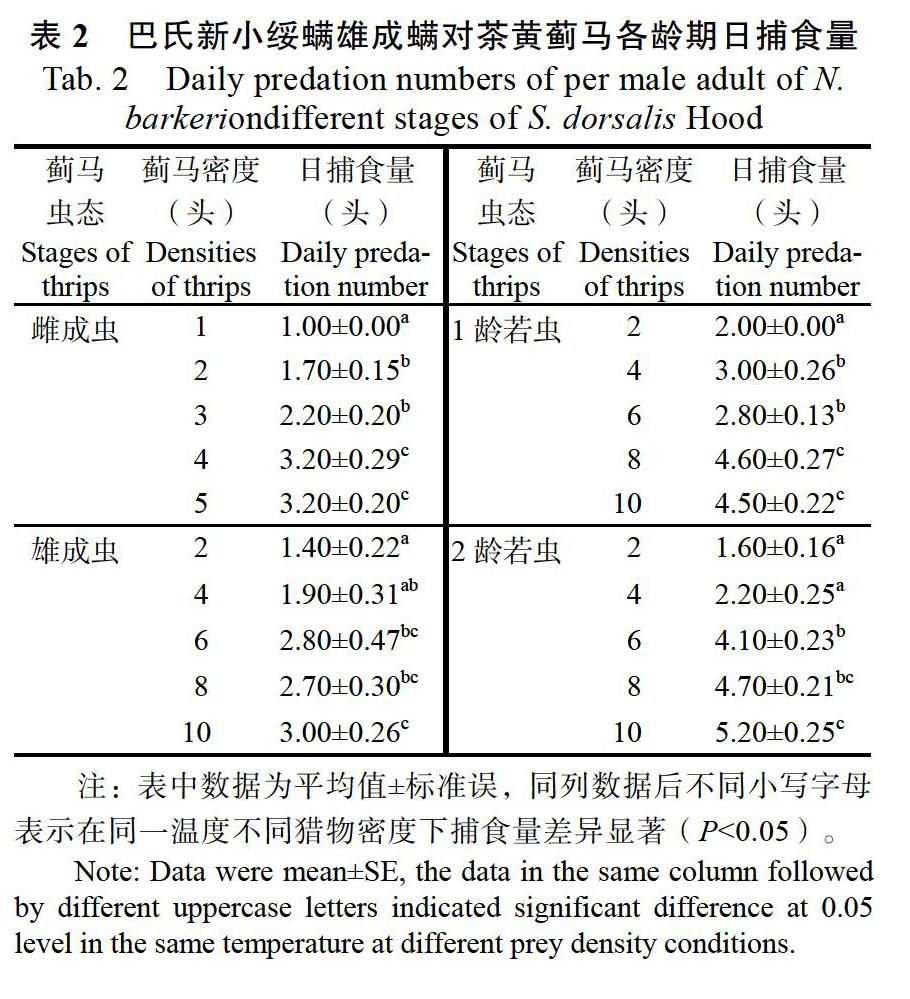
由表2可知,巴氏新小绥螨雄成螨对茶黄蓟马不同虫态的日捕食量均随猎物密度的增加而逐渐增加,当密度增加到一定程度时,其捕食量差异不显著。其中,对蓟马雌成虫的日捕食量较少,对1龄和2龄若虫的捕食量较多。
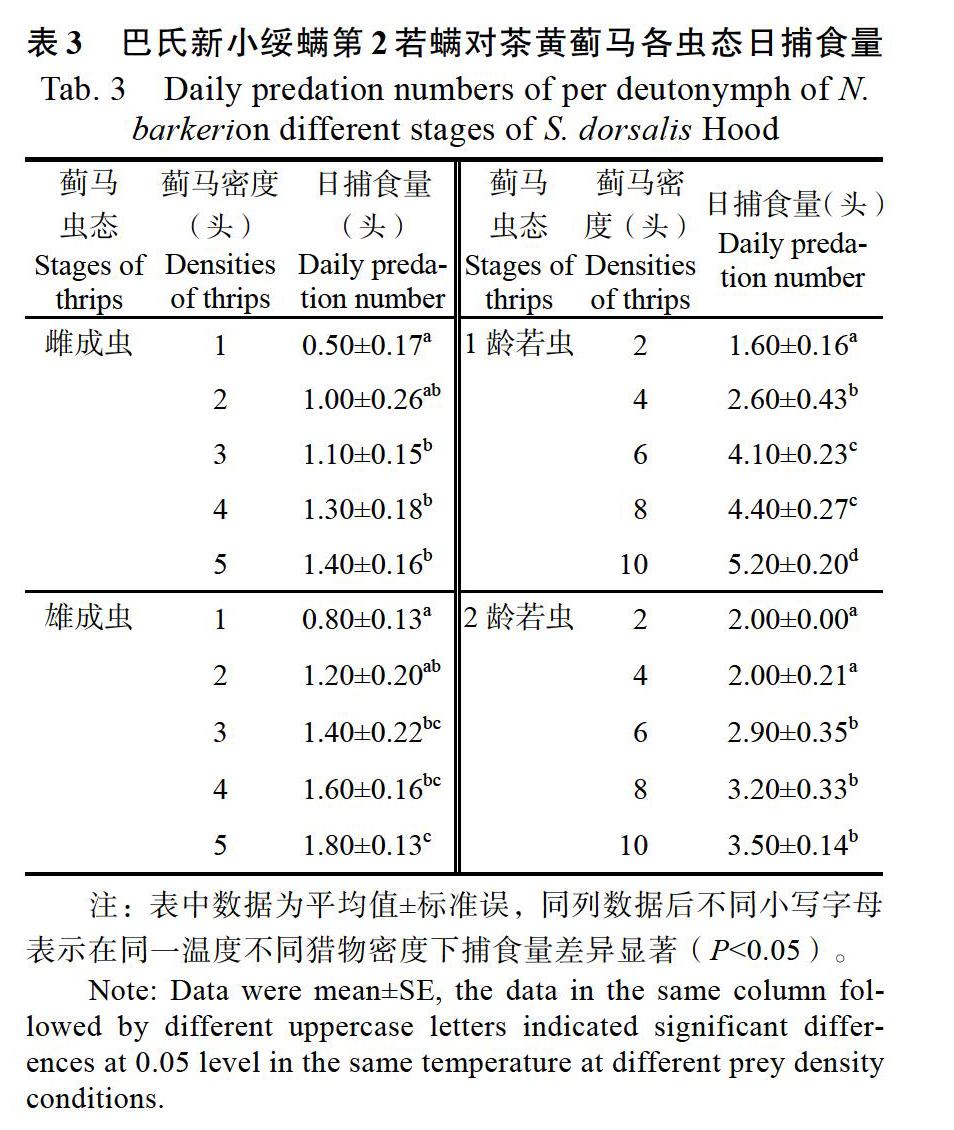
由表3可知,巴氏新小绥螨第2若螨对茶黄蓟马各虫态的日捕食量随猎物密度的增加而逐渐增加。其对蓟马的雌、雄成虫捕食量较少,对1龄蓟马若虫的日均捕食量较多。
2.2 巴氏新小绥螨对茶黄蓟马的功能反应
巴氏新小绥螨各螨态对茶黄蓟马各虫态的捕食作用采用Holling-Ⅱ型圆盘方程的拟合结果见表4。结果表明参数估计的R2值均大于或等于0.7023,拟合优度检验X2为0.0650~5.9757,X2 巴氏新小绥螨的雌、雄成螨和第2若螨对茶黄蓟马的不同虫态均具有较好的捕食效能。其中,雌成螨对蓟马1龄若虫的捕食效能最大,其次分别为蓟马雌成虫和2龄若虫,捕食效能分别为10.35、10.33和10.09头/d,对蓟马雄成虫的捕食效能和瞬时攻击率最小,对2龄若虫的瞬时攻击率最大,为1.0227。巴氏新小绥螨雄成螨对蓟马2龄若虫的捕食效能和瞬时攻击率最大,分别为12.48头/d和1.1208头/d,对1龄蓟马若虫的捕食效能和瞬时攻击率最小,分别为3.83头/d和0.6657。巴氏新小绥螨第2若螨对蓟马1龄若虫的捕食效能最大,为11.74头/d,其次为2龄若虫,对蓟马雌、雄成虫的捕食能力较低。总体来说,巴氏新小绥螨雌成螨和第2若螨对茶黄蓟马1龄若虫捕食效能最大,雄成螨对蓟马2龄若虫捕食效能最大。 2.3 巴氏新小绥螨对茶黄蓟马的寻找效应 不同螨态的巴氏新小绥螨对不同虫态的茶黄蓟马寻找效应不同,同一螨态巴氏新小绥螨对同一虫态的茶黄蓟马寻找效应随蓟马密度升高而下降。巴氏新小绥螨雌成螨对茶黄蓟马1龄若虫、2龄若虫和雌成虫寻找效应较好,其中对2龄蓟马若虫的寻找效应最好,对茶黄蓟马雄成虫寻找效应最差(图1)。 巴氏新小绥螨雄成螨对茶黄蓟马2龄若虫的寻找效应最好,其次是蓟马雌成虫,对蓟马雌成虫的寻找效应在密度較低时高于对1龄蓟马若虫的寻找效应,达到一定密度之后对1龄蓟马的寻找效应逐渐超过对蓟马雄成虫的寻找效应(图2)。巴氏新小绥螨第2若螨对茶黄蓟马1龄若虫的寻找效应明显高于2龄若虫,对蓟马雌成虫的寻找效应明显高于雄成虫(图3)。 3 讨论 本研究表明,巴氏新小绥螨适合作为芒果茶黄蓟马的天敌,且对芒果茶黄蓟马具有较好的控害潜能。 功能反应是评估捕食性天敌控害潜能的重要方法和途径。Holling-Ⅱ型圆盘方程模型是捕食者对其猎物的捕食功能最常见的形式。本研究表明,各螨态巴氏新小绥螨对芒果茶黄蓟马的不同虫态的功能反应属于Holling-Ⅱ型。这与前人所报道的巴氏新小绥螨对二斑叶螨Tetranychus urticae Koch、烟蓟马Thrips tabaci、腐食酪螨Tyrophagus putrescentiae、椭圆食粉螨Aleuroglyphus ovatus、截形叶螨Tetranychus truncatus Ehara[12-16]的功能反应拟合模型一致。其中,巴氏新小绥螨雌成螨对茶黄蓟马雄成虫的功能反应方程的R2值以及巴氏新小绥螨第2若螨对茶黄蓟马2龄若虫的功能反应方程的R2值均明显低于其他螨态的功能反应方程的R2值,表明其功能反应方程的拟合度相对其他功能反应的拟合度较差,然后通过X2检验来进一步验证方程的相关性,通过比较发现2组数据的X2值均明显小于规定临界值(df=49,=66.339),表明功能反应方程的拟合度符合要求。 本研究结果表明,不同螨态的巴氏新小绥螨对不同虫态的茶黄蓟马的捕食效能均有所差异。巴氏新小绥螨雌成螨对茶黄蓟马1龄若虫的捕食效能最大;巴氏新小绥螨雄成螨对茶黄蓟马2龄若虫的捕食效能最大;巴氏新小绥螨第2若螨对茶黄蓟马1龄若虫的捕食效能最高。通过比较最大日捕食量的差异,可以发现巴氏新小绥螨雌成螨对芒果茶黄蓟马1龄若虫最大日捕食量12.63头/d,略高于胡瓜钝绥螨对西花蓟马的最高日捕食量10头/d[17],明显低于斯氏钝绥螨对西花蓟马的日均捕食量达到最大18.15头/d[18],略低于巴氏新小绥螨对西花蓟马初孵若虫的最大日捕食量的14.47头/d[19],明显高于有益真绥螨、巴氏新小绥螨、加州新小绥螨、胡瓜新小绥螨和东方钝绥螨对西花蓟马1龄若虫的最大日捕食量的5.465、6.494、7.519、9.709和6.286头/d[20-21],表现出了较好的控害潜能。因此,可考虑巴氏新小绥螨作为芒果上茶黄蓟马重要天敌资源进行利用。 在觅食过程中,动物需要不断评估食物所含的能量和捕食食物所需要消耗的能量并作出决策[22],而且捕食者总是倾向于获得最大的净能量收益,即所获食物的粗能量(收益)和捕食消耗的能量(成本)之差[23]。本研究中巴氏新小绥螨对茶黄蓟马的若虫阶段相对于蓟马成虫表现出了较好的捕食选择性,这可能是由于蓟马成虫具翅有飞翔能力,对其进行捕食需要消耗更多的能量,综合衡量最佳效益,对无翅的蓟马若虫进行捕食效益更高。巴氏新小绥螨雌成虫对茶黄蓟马雌成虫、1龄若虫、2龄若虫均具有较好的捕食能力,这可能是由于在试验装置密闭条件下,茶黄蓟马雌成虫飞翔能力受到限制,在这种环境下捕食雌成虫对巴氏新小绥螨雌成螨可能在益耗比上相对有益,因此在室内试验观察表现具有较好的捕食功能。当捕食体型较大的被捕食者时,满足捕食者的数量将会较少[24]。巴氏新小绥螨第2若螨对茶黄蓟马1龄若虫捕食能力较好,这可能是由于1龄若虫个体较小,捕食单头1龄若虫所获得的能量较少,而巴氏新小绥螨雄成螨对茶黄蓟马2龄若虫捕食能力较好可能是由于其繁殖所需的能量较多,而且捕食行为将会带来一定量的能量损失[24],2龄若虫逃跑能力比蓟马成虫弱,提供的能量也比蓟马1龄若虫多,因此较为合适。由于在预蛹和蛹阶段,巴氏新小绥螨对其基本不捕食,且考虑到果园实际情况,预蛹和蛹是会落入地面,所以未对这2个阶段做捕食功能反应的试验。而且在不同温度下,巴氏新小绥螨对芒果茶黄蓟马的捕食情况也会呈现出差异性,后续试验将会考虑补充温度对其捕食情况的影响。另外,本研究仅仅是在室内对其控害能力进行评价,其控害效果还应结合田间释放效果来进行综合评价。 参考文献 周成刚, 李 健, 乔鲁芹, 等. 茶黄蓟马在银杏上的发生与危害[J]. 山东林业科技, 1994(4): 33 34. 尼章光. 云南芒果规范化栽培技术[M]. 昆明: 云南科技出版社, 2009. 劳有德, 韦文添. 芒果蓟马的为害与防治[J]. 广西热带农业, 2003(1): 16-17. 陈永森, 黄国弟, 覃 婵, 等. 芒果茶黄蓟马(Scirtothrips dorsalis Hood)田间发生动态及监测方法研究[J]. 南方农业学报, 2013, 44(10): 1646-1652. Moran M D, Hurd L E. Short-term responses to elevated predator densities: noncompetitive intraguild interactions and behavior[J]. Oecologia, 1994, 98(3-4): 269-273. Rosenheim J A, Kaya H K, Ehler L E, et al. Intraguild predation among biological-control agents: theory and evidence[J]. Biological Control, 1995, 5(3): 303-335. Hochberg M E. Consequences for host population levels of increasing natural enemy species richness in classical biological control[J]. American Naturalist, 1996, 147(2): 307-318. Janssen A, Pallini A, Venzon M, et al. Behaviour and indirect interactions in food webs of plant-inhabiting arthropods[M]//Bruin J, van der Geest L P S, Sabelis M W. Ecology and Evolution of the Acari. Dordrecht: Springer, 1999: 231-249. 孟瑞霞, 張青文, 刘小侠. 利用植绥螨防治烟粉虱的研究进展[J]. 昆虫知识, 2007(6): 798-803. 吴伟南. 中国动物志[M]. 北京: 科学出版社, 2009. Holling C S. Some characteristics of simple types of predation and parasitism[J]. The Canadian Entomologist, 1959, 91(7): 385-398. 尚素琴, 郑开福, 张新虎. 巴氏钝绥螨对二斑叶螨的捕食功能反应[J]. 植物保护学报, 2015, 42(3): 316-320. 黄建华, 罗任华, 秦文婧, 等. 巴氏钝绥螨对芦笋上烟蓟马捕食效能研究[J]. 中国生物防治学报, 2012, 28(3): 353-359. 王峻枫, 陈 军, 李桂亭, 等. 巴氏钝绥螨对腐食酪螨的捕食作用[J]. 安徽农业科学, 2013, 41(15): 6694-6696. 李朋新, 夏 斌, 舒 畅, 等. 巴氏钝绥螨对椭圆食粉螨的捕食效能[J]. 植物保护, 2008(3): 65-68. 崔晓宁, 张亚玲, 沈慧敏, 等. 巴氏钝绥螨对截形叶螨的捕食作用[J]. 植物保护学报, 2011, 38(6): 575-576. 郅军锐, 任顺祥. 胡瓜钝绥螨对西花蓟马的功能反应和数值反应(英文)[J]. 华南农业大学学报, 2006(3): 35- 38. 罗春萍, 户艳霞, 孙军伟, 等. 斯氏钝绥螨对西花蓟马的捕食功能反应[J]. 生物安全学报, 2018, 27(4): 274-278, 316. 尚素琴, 刘 平, 张新虎. 不同温度下巴氏新小绥螨对西花蓟马初孵若虫的捕食功能[J]. 植物保护, 2016, 42(3): 141-144. 马鹤娟. 加州新小绥螨对西花蓟马的控制作用初探[D]. 武汉: 华中农业大学, 2014. 郭建晗, 贾永红, 孟瑞霞, 等. 有益真绥螨对截形叶螨和西花蓟马的捕食功能反应[J]. 内蒙古农业科技, 2015, 43(3): 55-58. 黄雅梅. 基于ACT-R认知模型的界面设计要素及其交互方式研究[D]. 南京: 东南大学, 2015. 解 涵, 解玉浩. 鱼类摄食的经济学[J]. 河北渔业, 2003(6): 11-14. 朱新强. 基于能量流动的种群动力学模型[D]. 北京: 华北电力大学(北京), 2010.
