广西云耳菌株的遗传差异分析
2020-06-19吴圣进陈雪凤王灿琴苏启臣韦仕岩吴小建郎宁蓝桃菊
吴圣进 陈雪凤 王灿琴 苏启臣 韦仕岩 吴小建 郎宁 蓝桃菊


摘 要:為了解广西云耳菌株的遗传多样性,本研究对广西的10个百色云耳野生菌株和9个商品栽培菌株的菌丝和子实体性状进行测定,采用ISSR技术对其遗传差异进行分析,从而为广西云耳资源的合理利用提供依据。结果显示,19个菌株的菌丝培养特征和子实体外观性状都具有丰富的多样性;野生菌株与商品菌株间存在明显差异,其中前者多数菌株的菌丝培养期间色素发生早、子实体颜色较浅而呈半透明状、子实体质地柔软而滑嫩、咀嚼性低,后者菌丝培养期间色素发生晚或无色素、子实体颜色深而不透明、子实体质地较脆、咀嚼性高。ISSR分析结果显示,19个菌株的遗传相似系数在0.51~0.90之间,在相似系数为0.69时,可将它们划分为5个类群。其中,类群I和III全为野生菌株,遗传相似系数分别在0.69~0.74和0.69~0.77之间;类群II和IV全为栽培菌株,遗传相似系数分别在0.80~0.86和0.85~0.90之间;类群V仅野生菌株BY81。上述结果表明,广西云耳菌株具有丰富的遗传多样性,百色云耳野生菌株间的遗传差异较大,广西商品栽培的云耳菌株间的遗传差异较小;野生菌株与商品菌株间的遗传差异大,属于不同的生态类群;百色云耳是一类有待开发的宝贵云耳资源。
关键词:百色云耳;野生菌株;栽培菌株;ISSR;遗传差异
中图分类号:S646.6 文献标识码:A
Genetic Variation of Auricularia heimuer Strains in Guangxi
WU Shengjin, CHEN Xuefeng, WANG Canqin, SU Qichen, WEI Shiyan*, WU Xiaojian, LANG Ning,
LAN Taoju
Microbilogy Research Institute, Guangxi Academy of Agricultural Sciences, Nanning, Guangxi 530007, China
Abstract: In order to understand the genetic diversity and to provide a basis for the rational utilization of A. heimuer resource in Guangxi, the characters of mycelium and fruit body of ten wild strains from Baise and nine cultivars strains of A. heimuer were determined, and the genetic variation among of the strains was analyzed by the ISSR technique. The 19 strains exhibited high diversity in the characters of mycelium and fruit body. There were evident differences between the wild strains and the cultivar strains. The former mostly produced pigment early during mycelium culture, and the fruit body was light colored, semitranslucent, soft, tender, low chewiness in the textur, while the latter produced pigment at late stage of mycelium culture or no pigment, and the fruit body was dark colored, opaque, and springy, higher chewiness in the texture. ISSR analysis showed that the similarity coefficient was ranged from 0.51 to 0.90 among the 19 strains which could be divided into five groups at the similarity coefficient of 0.69. The strains in group I and group III were all wild strains, which had a similarity coefficient 0.69–0.74 and 0.69–0.77, respectively, the strains in group II and group IV were all cultivar strains which had a similarity coefficient 0.80–0.86 and 0.85–0.90, respectively, and the strain in the group V was only the wild strain BY81. All results suggested that A. heimuer strains in Guangxi possessed high genetic diversity. There were higher genetic variations among the wild strains from Baise than that among cultivar strains. There was also high genetic variation between the wild strains and cultivar strains, which should belong to different ecological groups. A. heimuer in Baise was a precious resource waiting to be exploited.
Keywords: Auricularia heimuer in Baise; wild strains; cultivar strains; ISSR; genetic variation
DOI: 10.3969/j.issn.1000-2561.2020.05.011
云耳是我国南方地区对黑木耳(Auricularia heimuer)的一种俗称,是我国四大食用菌栽培品种之一。我国云耳资源丰富、分布广泛,其中以秦巴山区、鄂西山区、云贵高原和东北黑龙江与吉林等地分布最多[1]。百色云耳指自然分布于广西百色地区及其周边地区的野生黑木耳,该地区位于云贵高原与南岭丘陵的过渡性地带,是广西云耳野生资源分布最多的地区,也是我国云耳的传统主产区[2]。
百色云耳是广西特有的黑木耳珍贵种质资源,与广西主要推广栽培的云耳品种在生物学特性、外观形态等方面有明显的差异[2]。百色云耳分布区域属于亚热带季风气候,云耳集中发生于3月下旬至5月中旬,属于中偏高温型品种[2-3];百色云耳以品质著称[5],耳片质地柔软,颜色以黄褐色为主[4]。研究表明,百色云耳在酯酶同工酶谱上存在丰富的遗传多样性,并蕴含许多的优异遗传性状[4],但其资源的评价和利用仍非常有限[7-8],其与广西商品栽培的云耳品种间的遗传差异尚未明确,其人工栽培等开发利用也非常有限。本研究采用分子标记技术(ISSR)进一步分析了百色云耳资源的遗传多样性,对比分析了百色云耳野生菌株与广西商品栽培云耳菌株的遗传差异,以期为百色云耳资源的挖掘利用提供依据。
1 材料与方法
1.1 材料
1.1.1 供试菌株 供试云耳菌株共19个,均保存于广西农业科学院微生物研究所,其编号、名称和来源见表1。
1.1.2 试剂与培养基 葡萄糖、琼脂粉均为国产分析纯,上海国药集团;6×Loading Buffer,2×Taq PCR MasterMix,GelRedTM购自生工生物工程(上海)股份有限公司。
PDA平板培养基:马铃薯(去皮)200 g、葡萄糖20 g、琼脂18 g、水1000 mL。
1.2 方法
1.2.1 菌丝培养特征和子实体性状观察 将供试云耳菌株用PDA平板培养基活化,再用灭菌好的打孔器取直径5 mm的菌种块,重新接种于新的PDA平板培养基,置于28 ℃下黑暗培养15 d,期间观察记录菌丝和菌落特征以及色素产生情况。
按照云耳常规袋料栽培方法[4]进行栽培出耳,待云耳耳片成熟后,观察子实体的颜色和透明度;将各菌株成熟耳片采收前喷水20 min,充分吸水后采收,选20名评委比较评价子实体手感的柔软程度,并比较评价沸水焯3 min再在凉水中冷却后的耳片口感和咀嚼爽脆情况。
1.2.2 菌株基因组总DNA抽提 将刚活化好的云耳菌丝接种于PDA培养基,28 ℃培养7 d,从培养基上刮取菌丝。采用Biospin真菌基因组DNA提取试剂盒(杭州博日科技有限公司)提取云耳菌株的基因组总DNA,用1.0%的琼脂糖电泳检测DNA质量。
1.2.3 ISSR分析 采用唐利华等[9]筛选出的编号为P3、P4、P5、P6、P9、P10、P16、P21、P22、P25、P26、P27和P32的13个ISSR引物,引物由生工生物工程(上海)股份有限公司合成。以供试云耳菌株的基因组DNA为模板进行PCR扩增(聚合酶链式反应),PCR反应体系参照韦仕岩等[10],PCR反应条件参照唐利华等[9]。PCR产物在2.0%琼脂糖凝胶中电泳,用稀释3300倍的GelRed溶液染色30 min,用凝胶成像系统观察拍照。
1.3 数据处理
选择稳定的ISSR扩增条带,记录各引物扩增条带的数据,有条带记为1,无条带记为0;采用
NTSYS-PC软件对云耳ISSR数据进行遗传相似系数分析,采用不加权成对群算术平均法(un-we ig hted pair group method with arithmetic means cluster analysis, UPGMA)進行云耳菌株的遗传相似性聚类分析,构建聚类树状图。
2 结果与分析
2.1 云耳菌株的主要外观性状
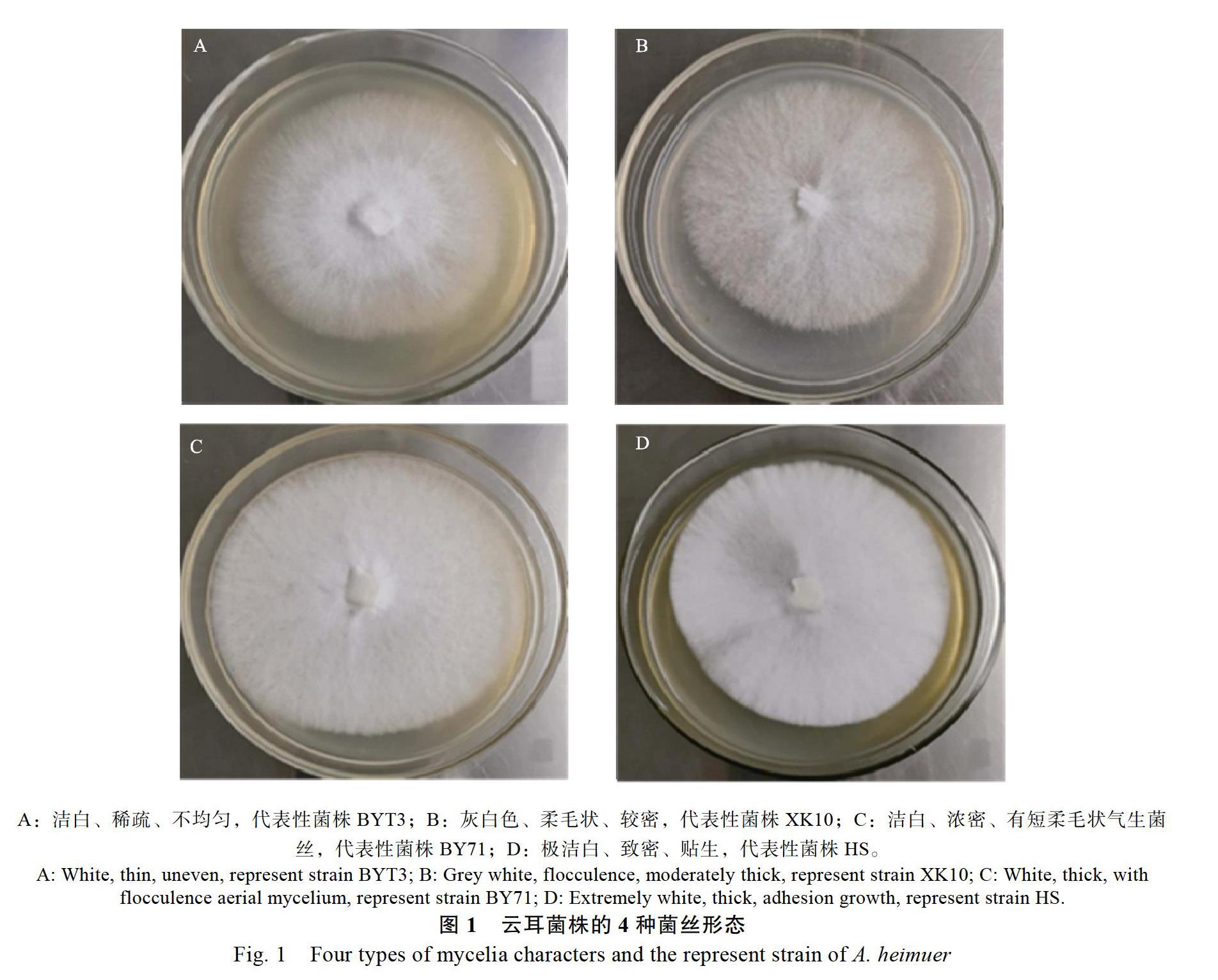
由表2可知,19个云耳菌株的菌丝培养特征大致可划分为4种类型:第1种类型为稀疏贴生为主,兼有少量柔毛状气生的菌丝(图1A);第2种为稀疏的柔毛状菌丝(图1B);第3种为浓密贴生为主,兼有部分柔毛状气生的菌丝(图1C);第4种为致密的贴生菌丝(图1D)。第1种类型的菌丝菌株4个,均为野生菌株;第2种类型菌株2个,均为商品菌株;第3种类型菌株9个(其中野生菌株6个,商品菌株3个);第4种类型菌株4个,均为商品菌株。在菌丝培养期间产生色素较早的菌株有4个,均为百色云耳野生菌株;其余菌株均在菌丝培养后期才产生色素,或者不产生色素。
由表2还可以看出,19个云耳菌株的子实体颜色表现较丰富的多样性,可分为半透明浅黄褐色(图2A)、半透明黄褐色(图2B)、深黄褐色(图2C)、黑褐色(图2D)、灰褐色(图2E)和半透明红褐色(图2F)6种类型。其中,子实体为半透明浅黄褐色至黄褐色的菌株有BY1、BY13、BY26、BY41、BY71和BYT3 6个野生菌株;深黄褐色的菌株有BY39、BY51和BY63 3个野生菌株及ME6、ME2和HME2 3个商品菌株;灰褐色至黑褐色菌株有H29、916、XK10、Au149、XK4和HS 6个商品菌株;子实体呈半透明红
褐色的菌株仅野生菌株BY81。由上述结果可知,多数野生菌株子实体颜色较浅,呈半透明状;商品菌株的子实体颜色均较深而不透明。此外,表2还显示,野生菌株的子实体质地柔软、口感滑嫩,而商品菌株的子实体柔软性稍差于百色云耳,口感偏脆。
2.2 云耳菌株ISSR结果分析
利用13个ISSR引物对19个云耳菌株进行PCR扩增,所有引物均能扩增出具有丰富多态性的清晰DNA条带,平均每个引物扩增的DNA条带数为23条,部分引物如P22扩增的DNA条带数达35条以上(表3、图3),总共获得DNA指纹谱带298条,其中多态性谱带288条,占总条带数的96.6%。研究结果表明,供试云耳菌株具有丰富的遗传多样性。
2.3 云耳菌株间亲缘关系分析
根据ISSR标记扩增结果进行遗传相似性分析,结果表明,19个云耳菌株的遗传相似系数在0.506~0.896之间,其中菌株XK4与HS的遗传相似度最大,而BY81与ME2的遗传相似度最小(表4)。
聚类分析表明,在相似系数为0.69时,可将19个云耳菌株划分为5个类群(图4)。其中,类群I和III为野生菌株,菌株数分别为3和6个,遗传相似系数分别在0.69~0.74和0.69~0.77之间;
类群II和IV为商品菌株,菌株数分别为4和5个,遗传相似系数分别在0.80~0.86和0.85~0.90之间;类群V为野生菌株BY81。研究结果表明,百色云耳野生菌株间的遗传差异较大,广西商品栽培的云耳菌株间的遗传差异较小;百色云耳野生菌株与商品云耳菌株分布聚在不同的类群,说明它们间的遗传差异大,亲缘关系较远。
3 讨论
形态学分析是研究食用菌遗传差异的传统方法,马庆芳等认为[11],以黑木耳子实体形态特征为依据可以鉴别不同菌株。黄艳芳等[2]认为,百色云耳以其子实体颜色鲜亮、质地柔软滑嫩等特征区别于推广栽培品种。本研究表明,广西云耳菌株的子实体外观性状具有丰富的多样性,百色云耳野生菌株与商品菌株间的外观性状差异明显(表2)。野生菌株普遍子实体颜色偏浅,半透明状,质地柔软,咀嚼性较低。但是,我们曾在不同栽培环境下观察到也有一些百色云耳野生菌株子实体颜色呈黑褐色[4, 12],与本研究结果不完全一致,这可能是由于栽培环境条件不同影响云耳子实体颜色所致。本研究是在相同环境条件下栽培不同云耳菌株,因此比较结果能反映它们的子实体颜色差别。本研究只能提供云耳子实体质地感官测定的定性结果,可能偏差较大,今后还有待借助质构分析仪进行定量测定[13]。
广西云耳菌株的菌丝培养特征多样,不同菌株的菌丝具有稀疏至浓密、气生和贴生等多种形态,不同菌株菌落产生色素的情况也不同(表2和图1)。百色云耳野生菌株与商品栽培云耳菌株相比,野生菌株中多数菌株菌落产生色素早,而商品菌株菌落产生色素晚,一些菌株甚至无色素产生。同时,商品菌株的菌丝相对更浓密、健壮。
ISSR分析结果表明,19个云耳菌株的遗传相似系数在0.506~0.896之间,说明广西云耳菌株具有丰富的遗传多样性,与酯酶同工酶分析的结论一致[4]。本研究直接采用唐利华等[9]筛选出的13个引物对19个云耳菌株进行ISSR扩增,结果表明,这些引物扩增的DNA條带数多、多态性高,说明这些引物完全适用于广西百色云耳和商品云耳的ISSR分析。亲缘关系分析表明(表4和图4),百色云耳野生菌株和商品栽培菌株分别归在不同的类群,前者遗传相似系数低于后者,说明野生菌株间的遗传多样性优于商品菌株,与前人研究报道一致[14-15]。与其他的研究报道相比[9, 15-17],广西商品栽培云耳菌株的遗传相似系数在0.80以上,遗传多样性更低,这可能是因为广西推广栽培的云耳品种少,菌株来源较窄,而且引进渠道多种多样,品种较混乱,可能有同物异名重复引进的现象。野生菌株和商品菌株分别归在不同的类群,说明二者的亲缘关系较远、遗传差异大,这可能是由于广西商品栽培的云耳菌株均来自外省,而广西本省的百色野生云耳虽有少量驯化栽培[7-8],但尚未得到商品推广栽培,还有待进一步开发利用。此外,野生菌株BY81与其他菌株的亲缘关系最远,形态差异最大,可能是不同的木耳品种。吴芳等[18]认为,黑木耳是共有5个种的复合种,传统使用的拉丁名A. auricular-judae是欧洲黑木耳种,而我国广泛栽培的中国黑木耳学名为A. heimuer,我国还分布有短毛木耳(A. villosula)、和美洲木耳(A. americana)。本研究对菌株BY81进行ITS序列比对,初步判断该菌株为黑木耳复合种中的短毛木耳(数据未给出),该菌株的分类地位还有待结合显微结构等形态学观察最终确定。
4 结论
广西云耳菌株在菌丝培养特征、子实体外观性状和DNA指纹图谱上均具有丰富的多样性,百色云耳野生菌株子实体颜色较浅而呈半透明状,质地更柔软、咀嚼性更低,从而在外观性状上明显区别于商品云耳菌株。野生菌株在分子水平上也与商品菌株有较大的遗传差异,两者亲缘关系较远,分属不同的类群。野生菌株间的遗传差异也较大,遗传多样性丰富,而商品菌株间的遗传差异较小。本研究结果可为百色云耳野生菌株资源的开发利用提供依据。
参考文献
余优森. 我国黑木耳气候生态条件分析与利用[J]. 中国农业气象, 1995, 16(5): 32-35.
黄艳芳, 李发盛, 黄雪星, 等. 广西野生黑木耳种质资源调查[J]. 中国食用菌, 2012, 36(6): 8-10.
李发盛, 李 槐, 何达崇, 等. 广西百色野生云耳生态环境调查[J]. 食用菌, 2008(5): 13-14.
吴圣进, 陈雪凤, 王灿琴, 等. 广西百色云耳野生菌株的遗传多样性分析[J]. 南方农业学报, 2018, 49(6): 1061-1067.
黄雄鹰. 近代右江地区对外贸易概述[J]. 广西民族研究, 1995(1): 12-18.
谢毅栋. 广西食用菌发展历程及对策[J]. 安徽农学通报(上半月刊), 2009, 15(21): 86-87, 129.
李发盛, 莫祺晖, 韦星明, 等. 云耳百云6号的生物学特性试验[J]. 食用菌, 2009, 31(3): 22-24.
王灿琴, 熊建文, 陈丽新. 五个野生云耳菌株的比较试验[J]. 食用菌, 2012, 34(2): 21, 24.
唐利华, 肖 扬, 边银丙. 中国黑木耳主要栽培菌株ISSR指纹分析及SCAR标记[J]. 菌物学报, 2008, 27(2): 243-251.
韦仕岩, 吴圣进, 汪 茜, 等. 草菇菌株的ISSR遗传差异分析[J]. 热带作物学报, 2013, 34(11): 2209-2213.
马庆芳, 孔祥辉, 戴肖东, 等. 东北地区黑木耳生产菌株栽培特性研究[J]. 食用菌学报, 2007, 14(4): 59-61, 91.
陈雪凤, 吴圣进, 王灿琴, 等. 23个广西野生云耳菌株的比较试验[J].食用菌, 2018, 40(3): 28-31.
么宏伟, 韩 越, 李元敬, 等. 不同培養基质和栽培方式对黑木耳感官品质影响[J]. 东北林业大学学报, 2017, 45(6): 51-56, 88.
刘华晶, 许修宏, 李春艳, 等. ISSR和ITS分子标记在黑龙江省野生黑木耳遗传多样性上的应用[J]. 东北农业大学学报, 2012, 43(8): 94-100.
张跃新, 律凤霞, 胡 伟, 等. 基于ITS序列分析黑龙江地区黑木耳菌株遗传多样性[J]. 中国林副特产, 2017, 147(2): 21-23.
任广明, 李滇华, 郭 兴, 等. 黑木耳栽培菌株亲缘关系的ISSR分析[J]. 东北林业大学学报, 2011, 39(5): 99-101.
李辉平, 黄晨阳, 陈 强, 等. 黑木耳栽培菌株的ISSR分析[J]. 园艺学报, 2007, 34(4): 935-940.
吴 芳, 戴玉成. 黑木耳复合群中种类学名说明[J]. 菌物学报, 2015, 34(4): 604-611.
