基于耐盐指数STTI的柳枝稷F2代群体耐盐性分析
2020-06-19刘一明孔丹宇刘小慧张训忠赵炳宇刘国道
刘一明 孔丹宇 刘小慧 张训忠 赵炳宇 刘国道

摘 要:雜交育种是植物耐盐新品种选育的重要途径之一,而对杂种F2代耐盐能力的综合评价至关重要。本研究以柳枝稷(Panicum virgatum)F2代的53个单株为研究对象,以亲本材料Alamo、Dacotah和F1代的DA90、DA120为参照,采用沙培法,在250 mmol/L NaCl的胁迫条件下进行耐盐性分析。在盐处理24 d后,测定电解质外渗率(EL)、相对含水量(RWC)、干重(DW)、净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs),通过对耐盐指数STTI进行主成分分析和隶属函数分析,结果表明:柳枝稷F2代的不同材料间耐盐能力存在显著差异。亲本Alamo的耐盐能力较高,亲本Dacotah的耐盐能力较差,F1代的DA90和DA120的耐盐能力中等,F2代的耐盐性有明显分离,表现在6份F2代材料的耐盐能力强于耐盐亲本Alamo,7份F2代材料的耐盐能力弱于不耐盐亲本Dacotah。主成分分析结果得到3个主成分,累计贡献率达87.47%。聚类分析将57份参试材料分为5类,分别代表不同耐盐程度的类群。隶属函数分析表明,F2代的39、45、27、48、35、53、7号耐盐性最强,该结果为柳枝稷耐盐相关性状的遗传图谱构建、QTL定位、柳枝稷耐盐育种奠定了基础。
关键词:柳枝稷;耐盐;STTI;F2代
中图分类号:S31 文献标识码:A
Salt Tolerance Analysis of F2 Generation Population of Switchgrass Based on Salt Tolerance Trait Index STTI
LIU Yiming1, KONG Danyu2, LIU Xiaohui3, ZHANG Xunzhong4, ZHAO Bingyu5*, LIU Guodao1*
1. Tropical Crop Genetic Resources Institute, Chinese Academy of Tropical Agriculture Sciences / Key Laboratory of Crop Gene Resources and Germplasm Enhancement in Southern China, Ministry of Agriculture & Rural Affairs, Danzhou, Hainan 571737, China; 2. College of Tropical Crops, Hainan University, Danzhou, Hainan 571737, China; 3. College of Agro-grassland Science, Nanjing Agricultural University, Nanjing, Jiangsu 210095, China; 4. Department of Crop and Soil Environmental Science, Virginia Tech, Blacksburg, Virginia 24061, USA; 5. Department of Horticulture, Virginia Tech, Blacksburg Virginia 24061, USA
Abstract: Cross-breeding is one of important ways to breed new varieties of plants, and it is essential to analyze the performance of hybrid F2 generation. In this study, 53 F2 generation plants of Panicum virgatum were used as the research material. The parents Alamo, Dacotah and F1 generation DA90 and DA120 were used as the reference. Salt tolerance performance was evaluated with sand culture method under 250 mmol/L NaCl stress conditions. The electrolyte leakage rate (EL), relative water content (RWC), dry weight (DW), net photosynthetic rate (Pn), transpiration rate (Tr), stomatal conductance (Gs) were measured after 24 days of salt treatment. Principal component analysis and membership function analysis of salt tolerance trait index (STTI) showed that there were significant differences in salt tolerance between different F2 plants. Parent Alamo was relatively salt tolerant, Dacotah was sensitive to salt, DA90 and DA120 had moderate salt tolerance, and the F2 generation had significant segregation in salt tolerance. Six F2 plants were better in salt tolerance than Alamo. Seven F2 plants were worse in salt tolerance than Dacotah. Three principal components with a cumulative contribution rate of 87.47% were selected by the principal component analysis. The 57 test plants were divided into five categories by cluster analysis. Membership function indicated that F2 generations 39, 45, 27, 48, 35, 53 and 7 had higher salt tolerance, which would build up a good basis for the construction of the genetic map of the salt tolerance-related traits, the QTL mapping and the salt-tolerant breeding of switchgrass.
Keywords: switchgrass (Panicum virgatum); salt tolerance; STTI; F2 generation
DOI: 10.3969/j.issn.1000-2561.2020.05.009
随着能源危机和环境污染的加剧,生物质燃料越发受到关注,发展生物质能源已成为应对气候变化、保障能源安全的重要手段[1]。柳枝稷(Panicum virgatum)为禾本科(Gramineae)黍属(Panicum)的多年生暖季型丛生禾草(C4),它原产于北美,在墨西哥、加拿大等地均有分布。1988年柳枝稷被引入我国,并逐步被广泛应用[2]。柳枝稷具有多年生、产量高、品质好、耐瘠薄、耐洪涝、耐干旱、抗病虫害、种植成本低等诸多优点,在观赏草中被广泛应用,同时也是一种优质牧草,目前作为模式能源植物进行研究[3]。柳枝稷主要有2种生态型:高地生态型和低地生态型。低地生态型多为4倍体(2n=4x=36),主要分布于南方潮湿地带,植株高大粗壮;高地生态型多为8倍体(2n=8x=72),主要分布在美国中部和北部地区,茎秆矮小[4]。柳枝稷可以作为盐碱化地区的植被种植,能够有效改善当地土壤环境,防止水土流失[5]。为避免“与粮争地”的现象发生,常用边际土地(盐碱、干旱等)种植柳枝稷。全球约有10亿hm2盐碱地,我国盐渍土地面积约1亿hm2[6],因此,可利用的边际土地资源十分丰富。
耐盐性属于多基因控制的数量性状[7],探討植物对盐分胁迫的适应机制、耐盐性的遗传改良一直是国内外育种工作者关注的热点,而利用杂交育种提高植物的耐盐性是植物新品种培育的有效手段。由于基因分离,F2代群体中会出现多种基因型个体,杂合体表现出较强的杂种优势[8]。柳枝稷种质资源较多,具有丰富的遗传多样性[9],利用柳枝稷进行杂交育种,从而改良柳枝稷的耐盐性,将会成为柳枝稷新品种培育的重要途径。目前,柳枝稷耐盐性的相关研究主要集中于耐盐综合评价、生理和分子机制方面。例如,Liu等[10]对30多份柳枝稷材料进行耐盐评价发现,低地生态型Alamo的抗盐性优于高地生态型Dacotah。此外,柳枝稷耐盐性表现出多种生理机制[11];在柳枝稷耐盐过程中,抗氧化酶(如CAT、POD、SOD)和microRNA发挥着重要作用[12-14];通过转基因手段,超量表达AANAT和HIOMT基因,能显著提高柳枝稷的耐盐性,转基因植株中的褪黑素含量显著升高[3];盐碱胁迫能够抑制柳枝稷种子的萌发[15]。目前,利用杂交手段提高柳枝稷耐盐性以及分析F2代柳枝稷群体的耐盐能力研究尚未见报道。
本研究以耐盐能力不同的柳枝稷亲本Alamo、Dacotah和二者杂交得到的F1代DA90、DA120以及53个F2代单株为材料进行耐盐能力分析,测定它们在盐胁迫下的生理指标变化情况,利用相关分析和主成分分析,了解柳枝稷F2代耐盐能力的表现,为柳枝稷的耐盐研究提供理论参考。
1 材料与方法
1.1 材料
以低地型Alamo为父本,以高地型Dacotah为母本,进行杂交,获得165个F1代单株,从F1代中选取不同单株进行杂交:单株5×49(母本),单株23×139(母本),单株90×120(母本),3组F1代杂交获得的F2代群体性状差异明显。本研究所用的53份F2代材料由90和120单株杂交所得,编号分别为1~53,亲本Alamo、Dacotah和F1代 DA90、DA120作为耐盐能力的参照(表1)。
1.2 方法
1.2.1 实验设计 采用沙培法,取柳枝稷材料进行分栽,种植于塑料盆(直径18 cm,高20 cm)中,每盆3株,每盆装4 kg沙(粒径0.1~1.0 mm)。定植后,在昼夜温度为(30±2)℃/(25±2)℃,光周期为12 h,光照强度为500 μmol/(m2·s),相对湿度为75%的温室内培养。当柳枝稷在温室内生长2个月后进行盐胁迫处理,盐处理选择250 mmol/L NaCl。盐处理浓度每12 h递增一次,依次从50 mmol/L增加至250 mmol/L,防止盐冲击效应。当盆内NaCl浓度达到250 mmol/L后,将塑料盆放在盛有Hoagland营养液的托盘中,营养液中的NaCl浓度为250 mmol/L,每天用蒸馏水补充蒸发的水分,使托盘内的盐浓度保持恒定。实验采用随机区组设计,4次重复。盐处理后的第24天进行指标测定。
1.2.2 指标测定及方法 (1)电解质外渗率。取50 mL离心管,加入20 mL去离子水,用电导率仪(SR60IC, VWR, Radnor, PA, USA)测定初始电导率E0;剪取充分伸展的柳枝稷上部成熟叶片,用蒸馏水洗净,吸干叶片上的水分,准确称取0.5 g叶片,剪成1 cm的小段,置于试管中振荡24 h,然后测定电导率E1。然后,将试管盖紧沸水浴30 min后取出,冷却至室温,摇匀后测定电导率E2。最后,计算电解质外渗率EL(Leaf Ele c trolyte leakage, %)[16],EL=(E1?E0)/(E2?E0)×100%。
(2)叶片相对含水量。剪取叶片约0.5 g,用天平称其鲜重(W1);然后将其浸入装有去离子水的试管中,封口浸泡24 h,称其饱和重(W2);然后,在105 ℃的烘箱中杀青30 min,85 ℃烘干至恒重,最后称其干重(W3)。计算公式为:叶片相对含水量=[(W1?W3)/(W2?W3)]×100%[17]。
(3)光合速率。采用美国Li-6400 XT光合仪测定叶片净光合速率[Pn, ?mol/(m2·s)]、气孔导度[Gs, mmol/(m2·s)]和蒸腾速率(Tr, mmol/mol)进行测定。光合测定参数设定:CO2浓度为400 ?mol/mol,流速为500 ?mol/s,光照强度为1000 ?mol/(m2·s)。测量时,叶片选上部的成熟叶片,每盆测3个叶片,取平均值。
(4)干重。在实验结束后,取所有植物植株,洗净植株根系表面的沙子,用吸水纸吸干表面残留的水分,置于烘箱中105 ℃杀青30 min,然后 85 ℃烘干至恒重,称其干重(DW)。
1.2.3 柳枝稷不同生态型的耐盐能力综合评价 采用耐盐指数(salt tolerance trait index, STTI)评价柳枝稷的耐盐性,STTI的计算公式为:STTI=各指标的处理值/各指标的对照值×100%[18]。利用模糊数学中的隶属函数对所有柳枝稷进行综合评价。
1.3 数据处理
采用SAS 8.1软件进行方差分析和最小显著差异性检验(LSD法),采用DPS 7.55软件进行相关分析、主成分分析和聚类分析,采用Excel 2016对F2代群体的耐盐相关指标进行频率分布分析。
2 结果与分析
2.1 盐胁迫对F2代53个柳枝稷电解质外渗率的影响
在盐处理(250 mmol/L NaCl)24 d后,所有实验材料的电解质外渗率明显高于对照,且各材料的电解质外渗率STTI存在显著差异(图1)。参照亲本Alamo和Dacotah的电解质外渗率STTI分别为133.60%和284.83%,F1代的DA90和DA120的电解质外渗率STTI分别为172.11%和226.56%,亲本Alamo的耐盐性显著高于Dacotah,
F1代DA90的耐盐性高于DA120,但差异不显著。F2代的14、2、19、7、45、27号的电解质外渗率STTI均低于126%,表现出较好的耐盐性,甚至高于耐盐亲本Alamo;F2代的26、49、4、36、33、30号的电解质外渗率STTI均高于317%,耐盐性相对敏感,比不耐盐亲本Dacotah表现还差。
2.2 盐胁迫对F2代53个柳枝稷相对含水量的影响
与对照相比,在盐处理24 d后,所有材料的相对含水量均呈下降趋势,且不同柳枝稷材料间相对含水量差异显著(图2)。柳枝稷亲本、F1代及53个F2代的相对含水量的趋势与电解质外渗率基本一致,耐盐能力依次为:Alamo>DA90>DA120> Dacotah,53个F2代材料的耐盐能力间隔分布其中,其中F2代的45、1、48、37、35号的耐盐性高于Alamo,相对含水量STTI均大于72.6%,而F2代的25、30、22、9、17号的耐盐能力比亲本Dacotah还要差,相对含水量STTI均小于56.2%。
2.3 盐胁迫对F2代53个柳枝稷净光合速率、气孔导度及蒸腾速率的影响
在250 mmol/L NaCl处理24 d后,所有柳枝稷材料的净光合速率、气孔导度、蒸腾速率都有所下降(图3,图4,图5)。57份材料间的净光合速率STTI、气孔导度STTI、蒸腾速率STTI均差异显著。根据3个光合相关指标的STTI可以看出,柳枝稷亲本Alamo的耐盐能力均优于F1代,F1代的耐盐能力均优于亲本Dacotah,而F1代DA90和DA120的耐盐能力在3个指标中的表现则有所区别,净光合速率和蒸腾速率的STTI显示DA90的耐盐能力优于DA120,而气孔导度STTI表明DA120的耐盐能力优于DA90。根据3个指标的STTI,F2代27、39、53、45号表现出较好的耐盐性,而F2代33、9、22、25、20号则表现出较敏感的耐盐能力。
2.4 盐胁迫对F2代53个柳枝稷干重的影響
在盐处理24 d后,干重分析结果表明,耐盐的柳枝稷材料的干重维持在较高水平,不耐盐的柳枝稷材料干重则明显降低(图6)。F2代48、Alamo、F2代45、53、32、11、7号表现出较高的耐盐能力,干重STTI均高于75.0%;F2代20、42、17、31、9、19号、Dacotah则表现出较差的耐盐能力,干重STTI均低于44.0%。
2.5 柳枝稷F2代盐胁迫下各指标STTI的相关性分析和主成分分析
相关性分析表明,电解质外渗率与相对含水量、净光合速率、蒸腾速率、气孔导度呈极显著负相关,相关系数依次为0.34、0.39、0.35、0.32(表2)。相对含水量、干重、光合速率、蒸腾速率、气孔导度之间都呈极显著正相关,其中净光合速率与气孔导度的相关系数最高,为0.88。
各指标STTI的主成分分析表明,主成分1、主成分2和主成分3的贡献率分别为64.95%、15.17%和7.35%,累计贡献率达87.47%(表3),主成分的公式为
PC1=?0.2298EL+0.4409RWC+0.3919DW+0.4614 Pn+0.4700Tr+0.4067Gs
PC2=0.9074EL+0.0597RWC+0.4127DW?0.0198Pn+0.0429Tr+0.0231Gs
PC3=0.0859EL?0.3383RWC?0.2005DW?0.2396Pn?0.0029Tr+0.8835Gs
以上3个主成分可以作为F2代柳枝稷耐盐能力评价的主因子。
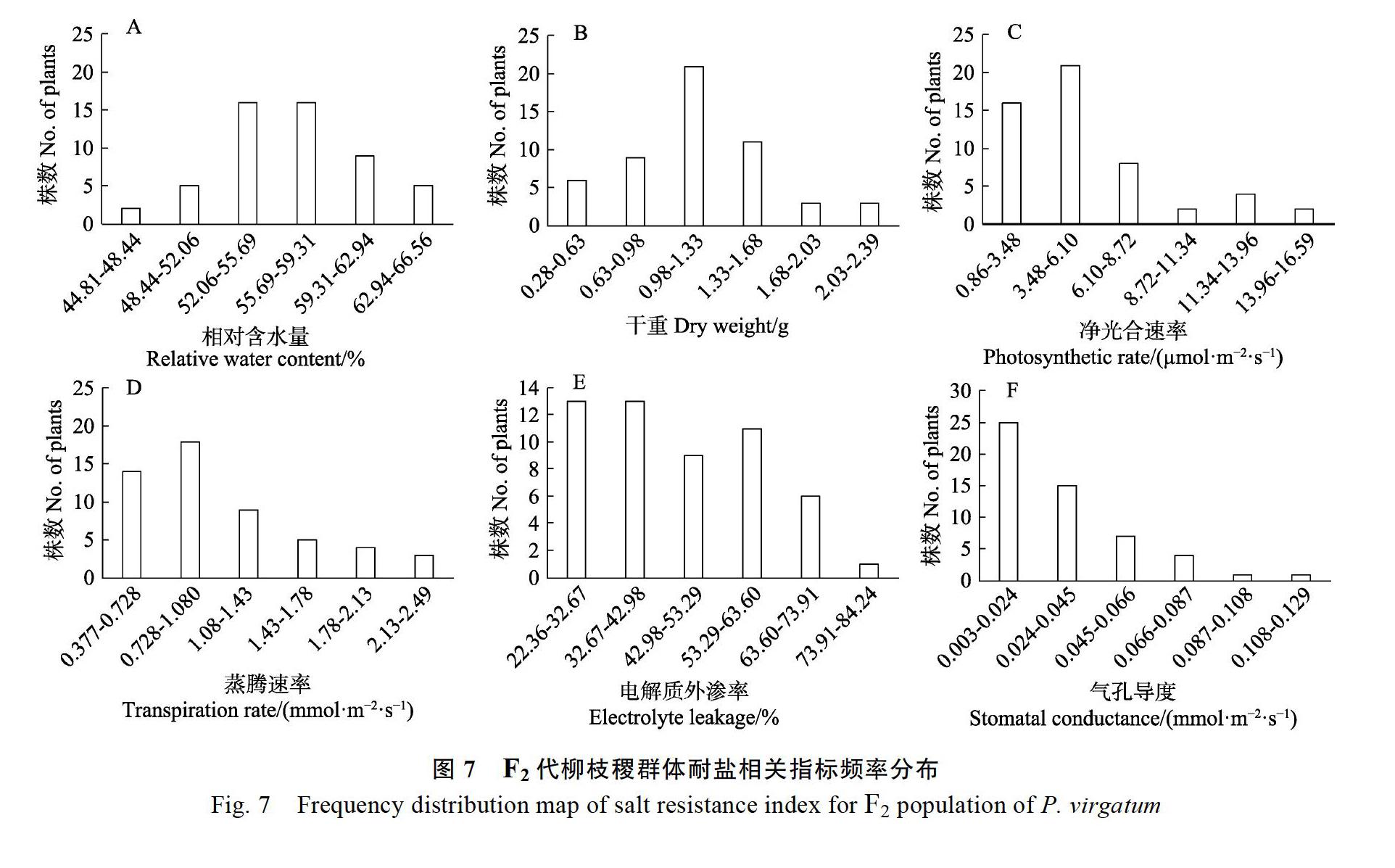
2.6 F2代柳枝稷群体耐盐相关指标的频率分布分析
在盐处理24 d后,分别对F2代53个柳枝稷材料的电解质外渗率、相对含水量、干重、净光合速率、蒸腾速率、气孔导度6个指标进行正态分布检验,分别以其分布范围为横坐标,以频数为纵坐标,采用Excel 2016软件对F2代群体的耐盐相关指标作图。结果表明,盐胁迫下F2代群体均出现较大变异,呈不同程度的连续分布,除电解质外渗率和气孔导度外,其他指标接近正态分布(图7)。
2.7 F2代柳枝稷群体的耐盐能力综合评价
Song L, Huang S C, Wise A, et al. A transcription factor hierarchy defines an environmental stress response network[J]. Science, 2016, 354(6312): aag1550.
陈全求, 詹先进, 蓝家样, 等. 棉花杂种F2代利用及发展研究进展[J]. 现代农业科技, 2014(23): 65-67, 70.
Liu Y M, Zhang X Z, Tran H, et al. Assessment of drought tolerance of 49 switchgrass (Panicum virgatum) genotypes using physiological and morphological parameters[J]. Biotechnology for Biofuels, 2015, 8(1): 152.
Kim J, Liu Y, Zhang X, et al. Analysis of salt-induced physiological and proline changes in 46 switchgrass (Panicum virgatum) lines indicates multiple response modes[J]. Plant Physiology and Biochemistry, 2016, 105: 203-212.
Liu Y, Zhang X, Miao J, et al. Evaluation of salinity tolerance and genetic diversity of thirty-three switchgrass (Panicum virgatum) populations[J]. BioEnergy Research, 2014, 7(4): 1329-1342.
Sun G, Stewart C N, Xiao P, et al. MicroRNA expression analysis in the cellulosic biofuel crop switchgrass (Panicum virgatum) under abiotic stress[J]. PLoS One, 2012, 7(3): e32017.
Wang Q, Wu C, Xie B, et al. Model analysing the antioxidant responses of leaves and roots of switchgrass to NaCl-salinity stress[J]. Plant Physiology and Biochemistry, 2012, 58: 288-296.
Xie F, Stewart C N, Taki F A, et al. High-throughput deep sequencing shows that microRNAs play important roles in switchgrass responses to drought and salinity stress[J]. Plant Biotechnology Journal, 2014, 12(3): 354-366.
Liu Y, Wang Q, Zhang Y, et al. Synergistic and antagonistic effects of salinity and pH on germination in switchgrass (Panicum virgatum L.)[J]. PloS One, 2014, 9(1): e85282.
Houimli S I M, Denden M, Mouhandes B D. Effects of 24-epibrassinolide on growth, chlorophyll, electrolyte leakage and proline by pepper plants under NaCl-stress[J]. EurAsian Journal of BioSciences, 2010, 4(1): 96-104.
Liu Y, Du H, Wang K, et al. Differential photosynthetic responses to salinity stress between two perennial grass species contrasting in salinity tolerance[J]. HortScience, 2011, 46(2): 311-16.
Tavakkoli E, Fatehi F, Rengasamy P, et al. A comparison of hydroponic and soil-based screening methods to identify salt tolerance in the field in barley[J]. Journal of Experimental Botany, 2012, 63(10): 3853-3867.
董荣书. 接种根瘤菌对柱花草耐盐性的影响及机理研究[D]. 海口: 海南大学, 2014.
崔彦茹. 利用选择育种群体进行水稻高产、抗旱和耐盐QTL定位[D]. 北京: 中国农业科学院, 2015.
邵 安. 利用小麥F2代(SR3 X JN17)群体进行盐胁迫相关主效QTL的SSR及EST-SSR定位[D]. 济南: 山东大学, 2010.
王喜濤. 黄瓜种质资源耐盐性评价及其遗传分析[D]. 哈尔滨: 东北农业大学, 2014.
张艳芳. 番茄特异PCR遗传连锁图谱的构建及苗期耐盐QTL定位[D]. 呼和浩特: 内蒙古农业大学, 2008.
龙卫华. 油菜发芽期耐盐评价、筛选与盐胁迫下根转录组分析[D]. 北京: 中国农业科学院, 2015.
孙 飞. 高粱杂交种耐盐性评价及耐盐生理机制研究[D]. 沈阳: 沈阳农业大学, 2018.
孙涌栋, 郝峰鸽, 徐 斌. 种子萌发期南瓜杂交品种耐盐评价[J]. 河南农业科学, 2009(10): 111-114.
Negr?o S, Schm?ckel S M, Tester M. Evaluating physiological responses of plants to salinity stress[J]. Annals of Botany, 2017, 119(1): 1-11.
Munns R, Tester M. Mechanisms of salinity tolerance[J]. Annual Review of Plant Biology. 2008, 59: 651-681.
Parida A K, Das A B. Salt tolerance and salinity effects on plants: a review[J]. Ecotoxicology and Environmental Safety, 2005, 60(3): 324-349.
Munir S, Siddiqi E H, Bhatti K H, et al. Assessment of inter-cultivar variations for salinity tolerance in winter radish (Raphanus sativus L.) using photosynthetic attributes as effective selection criteria[J]. World Applied Sciences Journal, 2013, 21(3): 384-388.
