基于叶绿体18S、CP12、CP15序列的西藏巨柏群体多态性遗传评价
2020-06-17张粟斌王子越林可心代靖娆段泽浩孟凡娟
张粟斌,雷 培,王子越,林可心,代靖娆,段泽浩,孟凡娟
(东北林业大学 生命科学学院,黑龙江 哈尔滨 150040)
巨柏Cupressusgigantea为柏科Cupressaceae柏木属Cupressus常绿乔木,又名雅鲁藏布江柏木,为西藏特有的珍稀树种,同时也是西藏特有的古树,寿命可达千年,材质优良,而且具有生态环境保护功能,是当地重要的造林、水土保持以及用材树种[1-4]。由于巨柏种群分布较为狭窄,天然更新困难,加之自然死亡以及人工砍伐等诸多因素的干扰,导致巨柏群体数量急剧减少,已成为我国濒危物种[5-6]。因此,了解巨柏遗传多样性特性,明确这个古老树种的遗传特性,对于揭示西藏巨柏的起源、保护和利用均具有重要意义。
叶绿体DNA(cpDNA)在多数被子植物中属母系遗传[7],其中非编码区变异丰富,对其进行测序分析可以广泛用于树木的遗传多样性和系统发育等研究[8-9]。此外,cpDNA用于树木起源与演化、种质资源保护等方面也较多[10-12]。本研究通过对67份西藏巨柏个体的叶绿体3个非编码区进行测序分析,明确西藏巨柏树种的起源与演化,从而为西藏巨柏种质资源保护和利用提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为雅鲁藏布江沿岸的巨柏种子,源自8个巨柏居群,共计67个个体,样品取样地信息见表1。将种子播种,获得的苗木叶片,直接进行冻存,存入-80 ℃超低温冰箱,以备后期叶片的总DNA提取。

表1 8个居群的样本信息
1.2 总DNA提取
采用改良后的CTAB法[10]提取巨柏叶片DNA,随后分别使用紫外光光度法和琼脂糖凝胶电泳法检测DNA浓度和纯度,取达到要求的DNA稀释至20 ng·μL-1,保存在-20 ℃冰箱用于PCR扩增。
1.3 PCR扩增和测序
通过查阅文献,本研究选择适合种内差异水平的 10 对非编码区的cpDNA通用引物(P02、CP12、CP15、18S、trnL-trnF、nadh K/C、matK、rbaL、rps4、trnQ-trnG)进行叶绿体PCR引物的初筛。所有PCR引物均由上海生物工程公司合成(表2)。
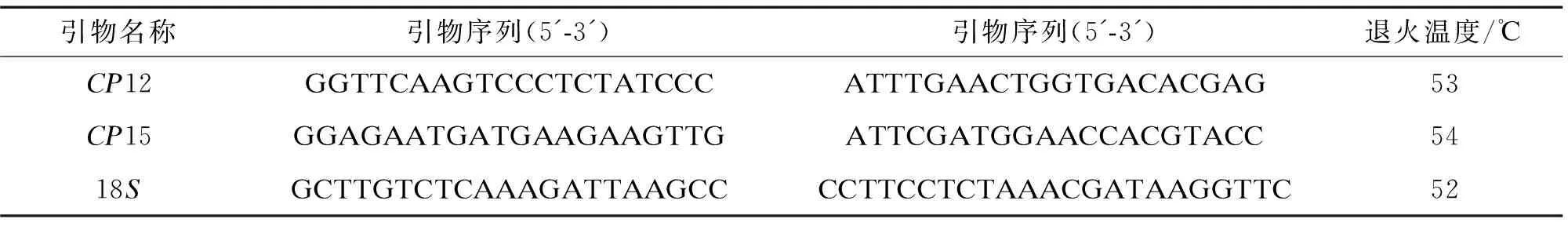
表2 试验采用cpDNA的引物信息
本试验采用单因素设计对反应体系进行优化,获得最佳的优化体系为:DNA模板2.5 μL、引物0.5 μL、10×PCR Buffer 2 μL、dNTP 2 μL、Taq DNA聚合酶0.2 μL、最后加入12.8 μL ddH2O,总体积为20 μL。反应程序:94 ℃预变性3 min;94 ℃变性30 s,65 ℃退火30 s,72 ℃延伸2 min,40个循环;72 ℃延伸10 min。选择凝胶成像清晰单一条带的PCR产物送吉林省库美生物科技有限公司进行双向测序。
1.4 数据分析
利用DNAstar和MEGA5.0软件对巨柏的叶绿体DNA测序片段进行拼接分析;利用DnaSP软件分析群体的单倍型多样性;通过Network软件构建叶绿体DNA单倍型间的中介网络关系图[13-14]。
2 结果与分析
2.1 序列特征
通过对测序的序列进行比对与对位排列后,CP12序列的长度为300 bp,CP15的序列长度为396 bp,18S的序列长度为1 000 bp。通过对3个序列的碱基分析发现:G/C含量丰富,在整个序列中所占比例为51.4%。图1为CP12扩增的所有样品的图谱。
2.2 单倍型特征
通过对3个序列进行比对,共发现变异位点4个,占序列全长的1.9%,其中18S区域存在2个变异位点共4个单倍型,CP12存在1个变异位点2个单倍型,CP15存在1个变异位点2个单倍型,各变异位点见表3。
2.3 遗传多样性
从表4可以看出:18S区域的单倍型多态性数据较高,其中k、Pi和Hd分别为0.754、0.008 5和0.569;非编码区CP12拥有较高的单倍型多样性方差(0.003 65)及单倍型多样性标准差(0.060)。

注:M为DNA Marker DL2000;1-67分别为67份巨柏材料。
图1 基于引物CP12的扩增图谱
利用T检验法分析分离位点数目与核苷酸多样性之间的关系。发现:当P<0.10时的D值均为正值,表示群体进化符合中性进化。T检验在扩增片段18S、CP12和CP15中均为正值(1.332 27、0.531 63和0.262 14),合并片段的结果也为正值,为0.967 80,根据差异显著性说明扩增的3个叶绿体DNA片段均遵循中性进化模型,进化过程中未经历过瓶颈效应。而且经过合并后的片段T检验结果也说明合并后的片段未经历瓶颈效应且遵循中性进化模型(表4)。
对不同巨柏群体进行AMOVA分析,群体间变异为14.2%,群体内变异为85.8%,可见群体内变异大,同时遗传分化系数Fst=0.224 80(P<0.001)。
2.4 单倍型的中介网络图构建
利用Network中介邻接网络算法和最大简约法构建巨柏不同居群的单倍型网络关系图(图2),共形成11个单倍型,其中出现频率最高的是H_1,其次为H_6,然后是H_7。其中群体4、群体8、群体15、群体14、群体13、群体12、群体11等7个群体共享单倍型H_1。由图2可以看出,单倍型3、单倍型4、单倍型9、单倍型11之间关系较近,但是单倍型分布概率较小,而且单倍型10和单倍型8与其它分支较远,说明由于长期进化,不同群体逐渐朝不同方向发展,导致群体11和群体14进化最远,而其它群体关系较近。

表4 叶绿体PCR片段多态性信息
注:H为单倍型数目;Vs为变异位点;Ps为简约信息位点;k为核酸差异平均数;Pi为核酸多态性;Hd为单倍型多态性;Vh为单倍型多态性的变异;Sh为单倍型多态性的标准变异。
3 讨论与结论
利用大量的基因组遗传多样性进行分析发现,通常叶绿体基因组的进化缓慢,且属于单向传递、不存在基因交换等特征,因此被广泛应用于遗传多样性分析。这主要是因为cpDNA作为细胞质DNA,在进化中不经历基因重组,导致受到的选择压力较小,可以反映植物在长期进化过程中的遗传与变异积累[15-16]。本研究针对8个群体进行叶绿体DNA(cpDNA)的遗传多样性分析,cpDNA的遗传多样性水平(Hd)为0.689,说明巨柏的遗传多样性较为丰富,对环境有较强的适应性,这与前人研究认为巨柏群体属于片段化的分布结果并不相同,而目前巨柏资源的衰退主要是诸多人为因素所导致的[17]。
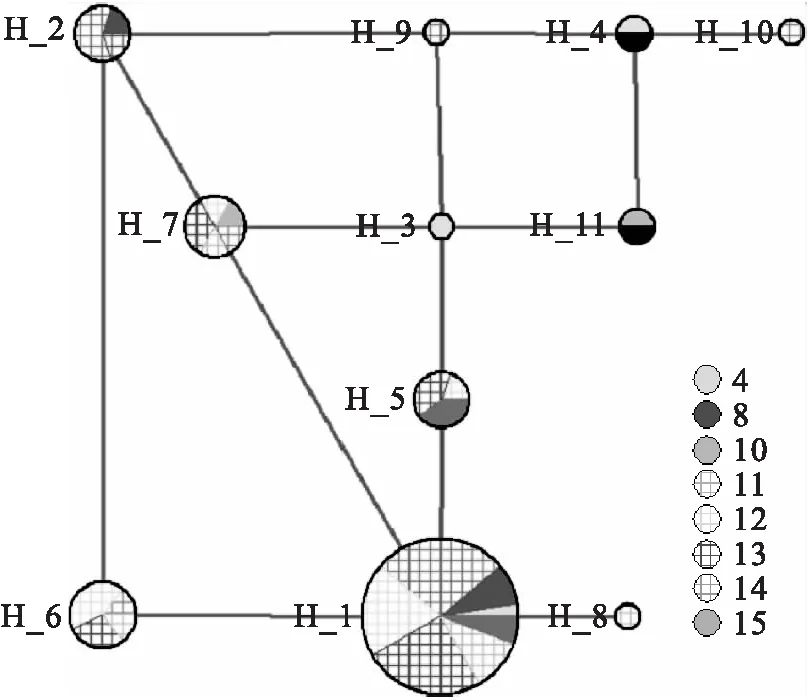
图2 基于叶绿体3种序列不同群体巨柏的中介邻接网络图
目前,主要采用叶绿体基因组的非编码区进行分析测定,如trnL-trnF、trnS-trnG和trnQ-trnG等,这主要是由于非编码区较为保守,可以对物种进行有效分析。但是在前期试验过程中,采用常用的叶绿体基因序列不能对巨柏进行有效扩增,扩增的多态性较小,因此,本研究所采用的序列并非常用的叶绿体基因序列,这可能是因为巨柏是较古老的树种,在基因组方面有其自身的特点,其原因还需要进一步研究。
通常采用T检验方法分析分离位点数目与核苷酸多样性之间的关系。当P<0.05或P<0.01的负D值说明群体进化过程中经历过瓶颈效应;当P>0.05时正D值,表示群体进化符合中性进化[18-19]。本研究发现:巨柏群体的D值为正值,说明巨柏的进化符合中性进化。同时对巨柏群体进行AMOVA分析,发现群体内变异比例较大(85.8%),个体的遗传变异对巨柏总遗传变异的贡献较大,群体间的遗传变异较小,也就是个体之间的遗传差异较大,因此对巨柏群体进行全面的个体保护措施,而不是采取群体保护措施。
综上分析,利用叶绿体序列可以有效揭示巨柏遗传多样性水平。同时根据巨柏整体遗传多样性水平高、种群内变异程度低的特点,采取对巨柏种子人工育苗等手段扩大种群数量,增加群体的遗传多样性;同时提高当地人民对巨柏濒危的认识,避免过度的人为采伐等破坏措施,宜采用就地保存的方式加以保护。
