水位梯度对狭叶香蒲光合作用的影响
2020-06-17周文昌史玉虎付甜庞宏东黄国伟
周文昌,史玉虎,付甜,庞宏东,黄国伟
(1.湖北省林业科学研究院, 湖北 武汉 430075;2.湖北洪湖湿地生态系统国家定位观测研究站, 湖北 洪湖 433200)
0 引言
水文、土壤和植被作为湿地生态系统三大组成要素[1],水文对湿地的存在起着决定性作用,决定了湿地景观、植被组成及其分布特征,严重影响着湿地植被生长[2].长江中游是我国暴雨频繁发生区域,每年的植被生长季,暴雨过程频繁,持续时间较长[3],这种暴雨将引起湿地水文波动,进而影响湿地植物生长[4].湖北省位于长江中下游地区,受东亚季风的影响,降水过程频繁,暴雨日数多,有“洪水走廊”之称,是中国易发生洪涝的地区之一,几乎每年都会遇到不同程度的洪涝灾害影响,特别是近几年,比如2016年6月30日至7月6日,24 h累积降雨量最大值约370 mm[5-6],超过特大暴雨划分等级250 mm的48%,导致内陆湖泊湿地蓄水量超载,引起内陆湿地内涝现象,导致湿地植物出现不同程度的水淹现象.这种特大暴雨事件严重影响植物的生长及其净光合速率,这是由于光合作用不仅是植物最重要的生理过程,而且是评价植物生长的标准之一,同时还是湿地植物生长繁殖与净化水体环境的能量来源[7-8].因此,研究水淹条件对湿地植物生长及光合作用具有重要意义,有助于人类认识湿地作为大气重要碳汇功能的变化规律.
本研究基于盆栽试验方法,以武汉九峰国家森林公园人工湿地种植的狭叶香蒲(Typhaangustifolia)植物为研究对象,将原状土壤+植物组合种植于试验桶中,采用不同水位深度控制,研究湿地挺水植物狭叶香蒲植物叶片光合作用能力对不同水位梯度的响应过程,为人类认识湿地植物对水位梯度变化下的湿地碳汇调节服务功能和湿地恢复策略提供理论依据.
1 研究区概况和研究方法
1.1 研究区概况本实验在武汉九峰国家森林公园(114°29′50″E,30°31′4″N)进行,采用2014年冬天整地与重建的人工湿地,湿地重建的底泥为库塘底泥,2015年3月购买水生植物种植于人工湿地中,植物包含美人蕉(CannaindicaL.)、狭叶香蒲、梭鱼草(Pontederiacordata)、鸢尾(Iristectorum),本研究以狭叶香蒲植物为研究对象.研究区内年平均气温16.3 ℃,极端高温41 ℃,极端低温-17.6 ℃,年日照数约1 600 h,无霜期240 d,年均降水量为1 200~1 400 mm,年均相对空气湿度79%.其森林植被主要以马尾松(PinusmassonianaLamb.)为主要常绿阔叶林,其次以优势树种枫香树(LiquidambarformosanaHance)、栓皮栎(QuercusvariabilisBl.)和马尾松为主要针阔混交林,森林覆盖率85%,有林地面积达483.52 hm2 [9].
1.2 研究方法2018年5月初,以研究区内人工沼泽湿地中狭叶香蒲为对象开展实验.生长季初期,狭叶香蒲萌生出约30~50 cm高度时,用铁锹将植株和原状土20 cm厚度一起移植至塑料圆桶内(规格直径35 cm×高度50 cm).然后,控制水位在5 cm、10 cm和20 cm深度,每个水位梯度形成3个重复.但是在将植株和土壤移植至圆桶中前,先在圆桶高度为25 cm、30 cm和40 cm处打2个小圆洞(对角处),以便维持水位在控制水位高度.为了让水位控制在同一水位,通过自来水向试验桶中加水,加水时间为每周1~2次,等待植物生长到旺盛期时(8月份),也就是生物量最大的时候,选择生长健康的植株开展测量不同水位梯度下的植物光合作用能力.
1.3 光合-光响应曲线2018年8月,选择晴朗无风的天气,在上午08:30~11:30,以美国生产的LI-6400XT光合系统测定植物生理指标,其仪器设置光合有效辐射(PAR)范围为0~2 000 μmol·m-2·s-1,梯度依次为2 000、1 500、1 300、1 200、1 000、800、500、250、200、150、120、60、30、15和0 μmol/(m2·s),二氧化碳(CO2)浓度控制在400 μmol/(m2·s),温度控制在30 ℃,湿度控制在70%~80%,选取生长健康的植株进行测量植物叶片的净光合速率,通过系统自动测量程序测定相应的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)等.
1.4 数据处理与分析数据处理通过Excel 2010和SPSS 18.0软件包,然后以光合有效辐射PAR为横轴、净光合速率Pn为纵轴绘制光响应曲线(Pn-PAR曲线)和最大光合速率(Pmax),并根据Farquhar生物化学模型非直角双曲线拟合曲线方程[10-11],方程式如下:
式中:Pn为净光合速率[μmol/(m2·s)],PAR为光强或光合有效辐射[μmol/(m2·s)],Pmax是植物最大净光合速率[μmol/(m2·s)],φ为表观量子效率(mol·mol-1),k为光响应曲线曲角,Rd为暗呼吸速率[μmol/(m2·s)].
通过对光响应曲线0~200 μmol/(m2·s)进行线性回归,Pn=0时,直线在x轴的截距为光补偿点(LCP,μmol/(m2·s));Pn=Pmax时,交点相应的x轴的数值为光饱和点(LSP,μmol/(m2·s))[12].
2 结果与分析
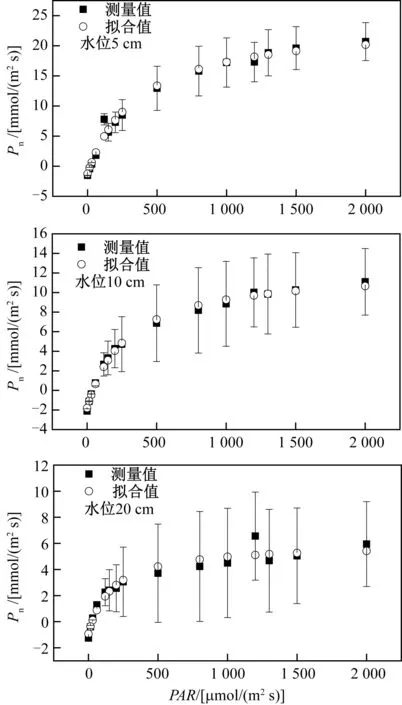
图1 不同水位条件下狭叶香蒲叶片的光合-光响应曲线
2.1 不同水位条件下狭叶香蒲光响应曲线的变化趋势3种水位条件下,光合有效辐射PAR位于0~250 μmol/(m2·s)之间时,植物净光合速率Pn几乎呈直线上升.当PAR超过250 μmol/(m2·s),Pn增速放缓,但仍然增大.当PAR超过500 μmol/(m2·s),除了水位20 cm时狭叶香蒲Pn变化幅度较小外,而水位为 5 cm和10 cm时,狭叶香蒲Pn仍继续增加(图1).但从图1中可知,狭叶香蒲生长于水位为 5 cm、10 cm和20 cm,存在水位高度越低,狭叶香蒲植物光合-光响应曲线越灵敏,也就是说狭叶香蒲种植在不同水位条件下,随着水位深度的增加,狭叶香蒲植物叶片净光合速率会出现明显的光抑制现象.3种水位深度下,狭叶香蒲植物叶片净光合速率Pn出现随水位深度的增加而净光合速率Pn越小.基于Farquhar生物化学模型拟合的光响应曲线,3种水位深度狭叶香蒲光响应曲线拟合的相关系数R2分别为0.988、0.996和0.947,拟合效果非常好(表1).
2.2 不同水位条件下狭叶香蒲光补偿点、光饱和点和最大净光合速率从表1中可知,狭叶香蒲水位深度为5 cm、10 cm和20 cm的光补偿点各自为18.94 μmol/(m2s)、48.32 μmol/(m2s)和27.16 μmol/(m2s);狭叶香蒲水位深度为5 cm、10 cm和20 cm的光饱和点各自为6 975.50 μmol/(m2s)、1 963.07 μmol/(m2s)和1 079.77 μmol/(m2s);狭叶香蒲水位深度为5 cm、10 cm和20 cm的最大净光合速率各自为25.430 μmol/(m2s)、14.241 μmol/(m2s)和6.886 μmol/(m2s).

表1 水位梯度下狭叶香蒲的光合-光响应曲线特征参数
2.3 不同水位梯度下狭叶香蒲植物净光合速率与生理生态因子的相关关系狭叶香蒲水位深度为5 cm、10 cm和20 cm条件下植物叶片净光合速率与气孔导度Gs、蒸腾速率Tr和光合有效辐射PAR均存在显著正相关关系,但与胞间CO2浓度Ci均存在显著负相关关系(表2).
3 讨论
从研究的结果来看,水位深度变化对狭叶香蒲植物叶片光合作用是有影响的,狭叶香蒲植物叶片的光合-光响应曲线的敏感度发生了变化.本研究的结果是当PAR在0~250 μmol/(m2s),Pn几乎呈直线上升,但当PAR在250~500 μmol/(m2s)之间时,净光合速率Pn增速放缓,但仍增大.当PAR>500 μmol/(m2s),水位20 cm时狭叶香蒲植物叶片净光合速率Pn变化幅度较小,而水位为5 cm和10 cm时,狭叶香蒲Pn仍继续增加;同时本研究表明狭叶香蒲植物叶片净光合速率随水位深度的增加,在光强增加后,出现了光抑制现象,并且植物叶片净光合速率也逐渐降低(图1).邓春暖等[2]研究表明芦苇(Phragmitescommunis)植物在不同淹水条件下,干湿交替处理下比持续淹水更有利于芦苇的生长,也就是说持续水淹降低了芦苇叶片的光合速率,干湿交替有利于提高芦苇产量,这与本研究的结果类似.同时,本研究结果也与张林海等[13]研究的闽江河口短叶茳芏(Cyperusmalaccensis)在短期(6个月)水淹的生物量高于中度水淹(15个月)的生物量类似.同时,这可以从本研究的饱和光强度下的狭叶香蒲植物叶片最大净光合速率可知,植物叶片的最大净光合速率出现随着水位深度增加,最大净光合速率呈降低趋势(表1).水位越高,狭叶香蒲植物叶片达到最大净光合速率越早,即光强越大,种植水位较高的狭叶香蒲植物净光合速率就越小.因此,本研究表明,在湿地恢复过程中,通过调控水位梯度,在滨岸带种植狭叶香蒲植物的湿地中,适当降低植物生长最佳时期的水位,有利于促进狭叶香蒲植物生长,产生更多生物量,进而固定大气中更多CO2,促使湿地碳汇能力增强.
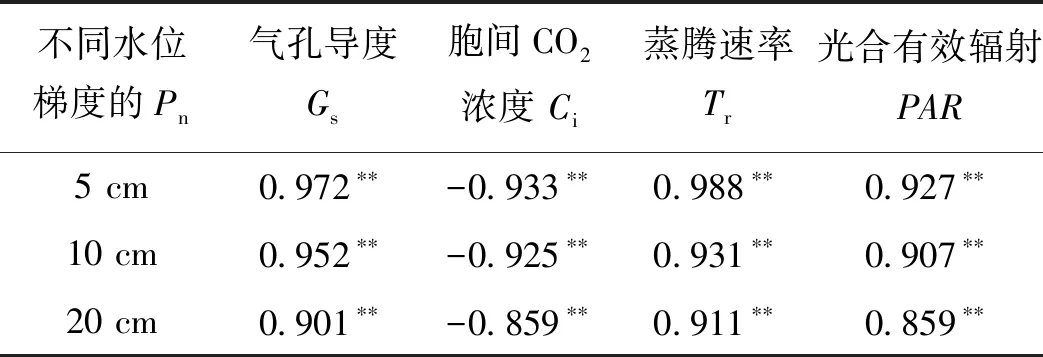
表2 不同水位梯度狭叶香蒲植物叶片净光合速率与生理生态因子的相关系数

图2 狭叶香蒲叶片光响应过程中净光合速率(Pn)与胞间CO2浓度(Ci)的关系
本研究的不同水位深度下狭叶香蒲植物叶片净光合速率与气孔导度Gs、蒸腾速率Tr和光合有效辐射PAR存在显著正相关关系,这与其他研究的结论吻合[14,15].但是在分析净光合速率与胞间CO2浓度的关系中,存在胞间CO2浓度起初较低时,随胞间CO2浓度增大,净光合速率呈增加趋势,然而胞间CO2浓度继续增大,净光合速率呈降低趋势(图2).这种情况详见陈根云等[16]针对此现象的解释,这种负相关的结果,是在不能使光合作用饱和的有限光下,光合速率随光合有效辐射增加而增高的理由是植物叶肉细胞光合活性增大,而不是胞间CO2浓度Ci降低;相反的是胞间CO2浓度Ci降低是由于叶肉细胞光合活性增大甚至光合速率增高.
光补偿点和光饱和点作为反映植物对光照的需求,各自代表的是植物对强光和弱光的利用能力[11,17].光补偿点较低、光饱和点较高的植物对光环境适应性强,而光补偿点高、光饱和点低的植物对光强的适应性较窄[17-18].通常光补偿点位于0~10 μmol/(m2s)的范围内,称为阴生植物,但是超过10 μmol/(m2s)后,为阳生植物[17,19].因此,狭叶香蒲植物为阳生植物.本研究表明水淹条件的变化,改变了狭叶香蒲植物光补偿点,但没有随着水位深度的增加而增加,而光补偿点出现递增或递减的趋势,水位为中间值(10 cm),光补偿点最大,这种现象有待于进一步研究.同时,光饱和点出现水位深度增加,狭叶香蒲植物叶片在最低水位时利用光能越强.本研究的光补偿点在18.94~48.32之间,这与张晓栋等[17]研究的菖蒲、鸢尾和美人蕉植物光补偿点的范围[11~47 μmol/(m2s)]吻合,但是本研究的光饱和点范围[1 079.77~6 975.50 μmol/(m2s)]远远大于张晓栋等[17]研究的菖蒲、鸢尾和美人蕉植物光饱和点范围[403~1 383 μmol/(m2s)],特别是水位深度在5 cm时的光饱和点[6 975.50 μmol/(m2s)],出现这种现象尚有待于深入研究,从5 cm深度水淹的狭叶香蒲植物光合-光响应曲线看出,光合有效辐射达到2 000 μmol/(m2s)时,植物叶片净光合速率已经达到20 μmol/(m2s),但是光强还需增加到近7 000 μmol/(m2s)时,即增加3.5倍左右,狭叶香蒲植物叶片净光合速率[25.43 μmol/(m2s)]才增加了27%.也就是说,光强超过2 000 μmol/(m2s)时,再增加,狭叶香蒲在5 cm这种水位高度下,净光合速率变化也是极小了,不过仍有待于下一步实验研究与探索.
4 结论
1)水淹条件的增加,影响了狭叶香蒲植物光合-光响应灵敏度,随着水深的增高,植物最大净光合速率降低,相当于光合能力降低.建议在湿地恢复过程中,在挺水植物狭叶香蒲生长最佳时期,适当降低水位有助于增加湿地碳汇功能,从而降低大气中CO2含量.
2)光合-光响应曲线表明,净光合速率与气孔导度、蒸腾速率和光合有效辐射呈正相关关系,但与胞间CO2浓度的关系有待商榷和探讨.
