高温与干旱胁迫对‘南林895杨’扦插苗生长和超微结构的影响*
2020-06-15杨佳骏吴永波张燕红
杨佳骏 吴永波 张燕红
(1.南京林业大学生物与环境学院 江苏省南方现代林业协同创新中心 南京 210037)
近年来,高温和干旱发生的频率和强度日益增加,植物正常生长发育受到危害(Rossati, 2017)。叶片是植物光合作用的主要场所,是树木发挥生态系统功能的基本单位(Wrightetal., 2004);叶片对环境胁迫敏感,其超微结构的变化能反映植物对环境胁迫的响应(Xuetal., 2006; Kratschetal., 2000)。非生物胁迫对植物生长的影响有多种形式,会引起一系列形态和生理变化。木本植物在生长过程中经常受高温与干旱的复合胁迫(Luoetal., 2008; Dai, 2013)。高温、干旱胁迫对林木光合作用性能、叶绿素荧光参数、抗氧化特性以及活性氧代谢等方面的影响已有大量研究(吴永波等, 2016; 叶波等, 2014; 李黎等, 2017; 张燕红等, 2017; Zandalinasetal., 2018),而复合胁迫下木本植物超微结构的变化却鲜有报道。
杨树(Populus)生长快、成材早、产量高,是我田中纬度平原地区种植面积最大的树种之一,在解决我国木材资源短缺及生态环境治理方面具有重要地位(方升佐, 2008),但极端高温与干旱严重抑制杨树的生长,从而影响其生态和经济效益(叶艳涛等, 2015)。目前,关于杨树应对高温与干旱胁迫的研究主要集中在生理特性、蛋白组学分析等方面(吴永波等, 2014),而关于其超微结构的变化研究却鲜有报道。本文通过研究在高温、干旱单一和复合胁迫下‘南林895杨’(Populus×euramericana‘Nanlin-895’)扦插苗生长以及叶片超微结构的变化,从形态学和细胞学角度探讨杨树生长发育对高温、干旱胁迫的响应机制,以期为杨树在高温、干旱地区的推广应用提供理论依据。
1 材料与方法
1.1 试验设计 试验材料为‘南林895杨’扦插苗,穗长20~23 cm,径粗2.0 cm。试验用盆的内径为30.0 cm,高为20.7 cm。供试土壤为黄棕壤,pH5.96,有机质含量15.14 g·kg-1,全氮含量0.83 g·kg-1,其中碱解氮含量15.79 mg·kg-1,全磷含量0.34 g·kg-1,其中有效磷含量1.34 mg·kg-1。在扦插3个月后将长势良好的杨树扦插苗放入3个不同温度的FYS-8智能人工气候室(3.5 m×2.5 m×3.0 m,高压钠灯光源,最大光合有效辐射为1 800 μmol·m-2s-1,最大光照度为80 000 lx,空气相对湿度为60%),白天温度分别为25 ℃(适宜温度T0)、35 ℃(中度高温T1)、40 ℃(重度高温T2),夜间温度均为25 ℃。设置3个水分梯度,土壤含水率分别为田间持水量的70%(适宜水分D0)、55%(中度干旱D1)、40%(重度干旱D2),即有T0D0(对照)、T0D1、T0D2、T1D0、T1D1、T1D2、T2D0、T2D1、T2D2共9个处理。每个处理设4个重复,共36盆。试验期间每天上午6:00,采用称量与TR200水分测定仪相结合的方法,对每盆植物进行称量并补充水分,确保土壤含水率在控制处理下。同时利用气候室模拟自然环境,调节光照量,于每天6:00打开光照、18:00关闭光照。胁迫持续7周后,在第50天测定指标。
1.2 试验方法 1) 形态指标测量 利用卷尺和游标卡尺分别测定扦插苗的株高、地径。用自封袋自上而下收集各处理的杨树苗生长完全展开的叶片5片,用扫描仪(Cano,Japan)进行扫描,将扫描图片用ArcGIS软件测量叶片的周长和面积,叶缘分裂程度=叶周长/叶面积。
2) 生物量测量 将不同处理下各4株杨树扦插苗用蒸馏水冲洗干净,吸水纸吸干,将根、茎、叶分开后分别置于信封袋中,放置于105 ℃烘箱中杀青15 min,75 ℃烘干至恒质量,测定各部分生物量。生长指标:根茎比=地上生物量/地下生物量、叶分配指数=叶生物量/总生物量,茎分配指数=茎生物量/总生物量,根分配指数=根生物量/总生物量。
3) 叶片超微结构观察 分别取各处理的扦插苗完全展开的功能叶片(自上往下数第3片),用刀片切得叶片中部大小1 mm×1 mm的叶片(不含叶脉),迅速放入浓度为2.5%的戊二醛固定液中,用针筒抽去空气使叶片沉入固定液(真空状态),在戊二醛中固定24 h以上,用0.1 mol·L-1的磷酸缓冲液漂洗3次后,用 1%锇酸在4 ℃下固定过夜,磷酸缓冲液漂洗; 再用乙醇系列脱水,丙酮过渡,Epon812树脂渗透与包埋。LKB-V超薄切片机切片; 醋酸双氧铀以及柠檬酸铅双染; H-600型透射电镜下观察并照相。
1.3 数据处理与分析 利用Microsoft office 2010进行数据处理分析; 采用SPSS 17.0进行方差分析,Duncan新复极差法进行显著性检验。
2 结果与分析
2.1 高温和干旱胁迫对‘南林895杨’扦插苗形态特征的影响 株高和地径作为生长指标可直观反映胁迫下的植株生长状况。由表1可知,高温、干旱单一和复合胁迫使株高和地径均有不同程度下降,高温胁迫下株高和地径间差异显著(P<0.05),表现为T2D0
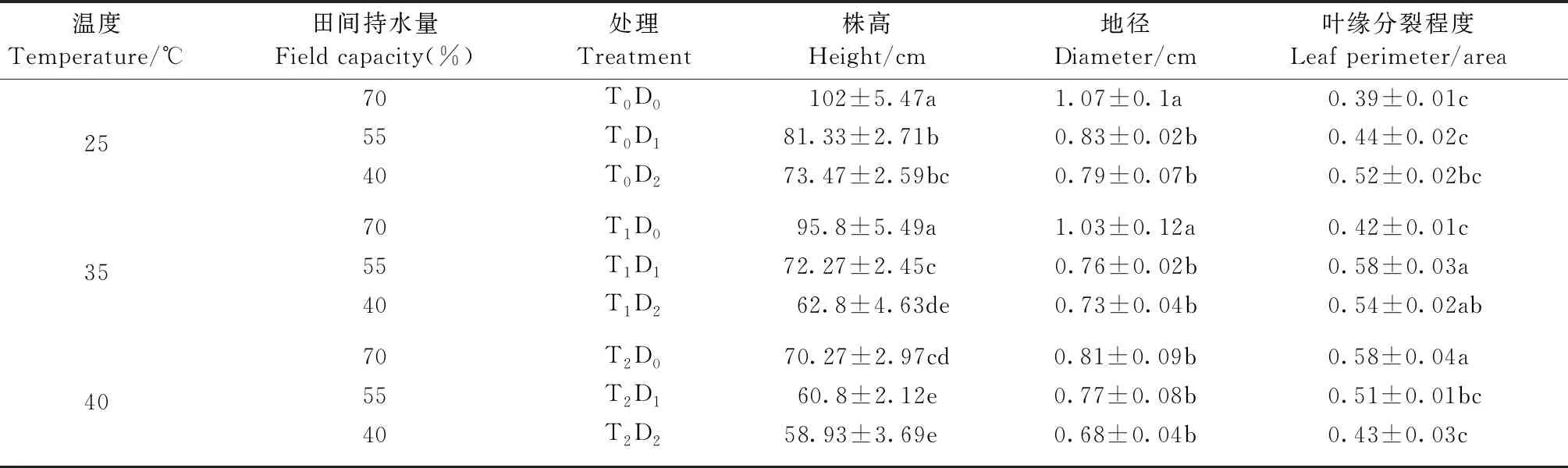
表1 高温和干旱胁迫对‘南林895杨’幼苗生长的影响①
①同列数字后不同小写字母表示处理间差异显著(P<0.05)。 下同。Different lowercase letters in the same column indicate significant differences between treatments (P<0.05). The same below.
叶缘分裂程度因高温、干旱单一和复合胁迫而增大。在高温或干旱单因子胁迫下,叶缘分裂程度随胁迫强度增加而增加,T1D0、T2D0、T0D1和T0D2分别比对照增加7.3%、49%、12.5%和33.5%,但仅T2D0处理与对照差异显著(P<0.05)。T1D1、T1D2复合胁迫下的叶缘分裂程度显著高于温度单一胁迫处理(P<0.05),分别较对照增加48%和38%。
2.2 高温和干旱胁迫对‘南林895杨’扦插苗生物量分配特征的影响 由表2可知,随单一和复合胁迫程度加剧,地上生物量、茎生物量、叶生物量均不同程度下降。高温单因子胁迫下,各处理间的地上生物量、茎生物量差异显著(P<0.05); 干旱单因子胁迫下,各处理间无显著差异(P>0.05); 复合胁迫处理下,地上生物量、茎生物量、叶生物量均低于单一胁迫处理。
高温、干旱单一和复合胁迫使根生物量下降。高温胁迫下,各处理间差异显著(P<0.05),T1D0、T2D0处理分别较对照下降9.4%、39.4%,干旱胁迫下,T0D1、T0D2处理分别较对照下降12.7%、24.8%,其中T0D2处理与对照差异显著(P<0.05),复合胁迫T1D1、T2D1、T2D2处理下,根生物量均低于单因子胁迫处理,分别较对照下降17.2%、45.7%、47.4%。T1D2处理的根生物量高于T0D2单一胁迫处理,较对照下降15%(表2)。
高温、干旱单一和复合胁迫下,随着胁迫程度加剧,扦插苗总生物量下降显著(P<0.05)。高温胁迫处理间差异显著(P<0.05),T1D0、T2D0处理分别较对照下降13.6%、52.1%,其中T1D0处理与对照差异不显著(P>0.05); 干旱胁迫T0D1、T0D2处理的总生物量分别较对照下降24%、38.1%,与对照差异显著(P<0.05); 复合胁迫T1D1、T1D2、T2D1、T2D2处理下总生物量低于单一胁迫处理,分别较对照下降25.9%、35.7%、59.5%、63.0%(表2)。
高温、干旱单一和复合胁迫下,杨树幼苗根茎比升高。高温胁迫T1D0、T2D0处理分别较对照升高6.4%、38%,两者间差异显著(P<0.05); 干旱胁迫T0D1、T0D2处理分别较对照升高21%、27.9%,两处理之间无显著差异(P>0.05),表明重度高温胁迫对根茎比的影响大于干旱胁迫。中度高温胁迫对根茎比无显著影响(P>0.05); 复合胁迫下根茎比上升幅度大于单一胁迫,T1D1、T1D2、T2D1、T2D2处理分别较对照升高13.9%、48.4%、48.4%、64.5%,但与单一胁迫差异不显著(P>0.05)(表2)。
2.3 高温和干旱胁迫对‘南林895杨’扦插苗叶肉细胞超微结构的影响 1)正常条件下叶肉细胞的超微结构 正常温度和水分条件下的扦插苗叶肉细胞超微结构如图1。叶肉细胞结构完整,细胞内含有中央大液泡,液泡膜光滑清晰(图1A、B)。细胞核呈球形,核被膜清晰,染色质均匀分布(图1C)。叶绿体呈椭圆形或卵圆形,结构完整,淀粉粒分布均匀(图1D)。叶绿体基粒片层清晰、排列紧密未出现解体现象,叶绿体内含有少量的嗜锇体(图1E)。

表2 高温和干旱胁迫对杨树苗器官生物量分配的影响

图1 正常温度和水分条件下的‘南林895’扦插苗叶肉细胞的超微结构

图2 高温或干旱单一胁迫下‘南林895杨’扦插苗叶肉细胞的超微结构
2) 高温或干旱单一胁迫下叶肉细胞超微结构的变化 中度高温T1D0处理下,杨树扦插苗叶肉细胞结构正常,排列紧密,未发生质壁分离现象(图2A)。细胞核结构完整,核膜清晰(图2B)。叶绿体排列规则紧贴细胞壁,略有膨胀,但不明显(图2C)。淀粉粒含量未发生太大变化。叶绿体内嗜锇颗粒变大(图2A、C),基粒片层排列整齐稍有扩张(图2D)。重度高温T2D0处理下,细胞结构正常,个别叶肉细胞出现质壁分离的现象(图2E箭头)。细胞核结构正常,核膜清晰(图2F)。叶绿体膨大变圆,部分发生消解。淀粉颗粒减少,嗜锇体数量大量增加(图2G)。基粒片层扩张,出现空泡(图2H)。中度干旱T0D1处理下,杨树叶肉细胞结构完整,排列整齐(图2I)。细胞核核膜清晰(图2J)。叶绿体结构完整,数量减少且有膨大变圆趋势。淀粉粒含量减少,嗜锇颗粒增多(图2K),片层出现扩张现象(图2L)。重度干旱T0D2处理下,杨树叶肉细胞结构完整,细胞形状呈狭长型(图2M)。核膜部分边界模糊(图2M)。叶绿体膨大变圆,位于细胞中间。淀粉颗粒减少(图2O),嗜锇体变大,片层扩张明显,部分消融(图2P)。
高温胁迫与干旱胁迫对杨树叶肉细胞超微结构的影响有许多异同点。超微结构显示,高温胁迫对细胞器膜结构的影响要比干旱胁迫显著,而细胞形状和叶绿体数量对干旱胁迫有明显响应。
3) 高温和干旱复合胁迫下叶肉细胞超微结构的变化 T1D1处理下,细胞结构完整。叶绿体数量较高温胁迫减少,膜结构部分消失(图3A)。细胞壁外表面出现少量的黑色致密物质(图3B、C,箭头)。核膜结构不清晰(图3B)。嗜锇颗粒数量较单一胁迫增加(图3C),叶绿体片层扩张明显(图3D)。T1D2处理下,细胞质壁分离严重,少量细胞形态扭曲(图3E),细胞壁外表面有少量黑色致密物质(图3F、G箭头),核膜消失,细胞核开始解体(图3F)。叶绿体膨大变圆边界模糊,淀粉粒消失,嗜锇颗粒数量较单一胁迫明显增加(图3G),叶绿体片层严重扩张,部分消融(图3H)。T2D1处理下,叶肉细胞严重扭曲(图3I),叶绿体膨大变圆边界模糊,质壁分离现象严重(图3I、J箭头)。细胞核形状不规则,核质凝集(图3K)。叶绿体膜结构不清晰,有内含物流出(图3K)。淀粉颗粒较单一胁迫减少,嗜锇颗粒较单一胁迫增大增多(图3K、L)。叶绿体片层模糊,消融严重(图3L)。T2D2处理下细胞结构扭曲(图3M),细胞出现严重的质壁分离现象(图3M箭头)。细胞内膜系统大部消失。细胞核消融严重,胞质外流(图3N)。叶绿体数量减少,消融严重基质外流(图3M)。淀粉颗粒数量减少,嗜锇颗粒增加(图3O)。细胞壁外表面有黑色致密物质(图3O箭头),叶绿体片层扩张甚至扭曲,空泡化严重(图3P)。
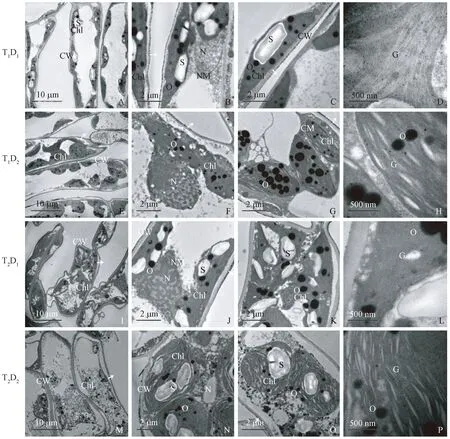
图3 高温和干旱复合胁迫对‘南林895杨’扦插苗叶肉细胞的超微结构的影响
3 讨论
3.1 高温和干旱胁迫对杨树形态和生长的影响 高温、干旱胁迫下的‘南林895杨’扦插苗的株高、地径均呈下降趋势,根、茎、叶的生长都受到抑制。虽然扦插苗对中度高温干旱表现出一定抗性,但重度高温干旱胁迫对扦插苗生长抑制较为明显。复合胁迫对生长的危害更严重,在高温条件下植物需要通过提高蒸腾来降低叶片温度以维持其正常功能,同时要避免干旱条件下蒸腾过快加速土壤水分消耗。因此,复合胁迫下生长的幼苗需发育出大小适宜的叶片来平衡水分利用与叶表温度。高温环境中生长的植物多具有大的叶片,通过增大蒸发面积达到快速降低叶表温度的目的,而干旱区的植物叶片偏小则是长期适应水分亏缺的结果。叶形在高温干旱的条件下对叶表的降温作用明显,因为在干旱条件下蒸腾的降温作用受到抑制,植物更多地依赖叶形改变来降低边界层阻力从而增加叶片散热(李永华等, 2018)。叶缘分裂程度(周长面积比)可作为叶片边界层阻力的特征值(Roth-Nebelsick, 2001)。与对照相比,单因素胁迫和复合胁迫下的叶缘分裂程度均增加,说明在叶片变小过程中的周长降幅小于面积降幅。在干旱区,叶长的优先变化有利于周长的变化,宽度的优先变化有利于叶面积的变化(李永华等, 2012),说明杨树叶片在胁迫条件下趋向变窄。通过模拟叶片的热成像可知,小而窄的叶片边界层阻力小,因此温度较低且降温更快(李永华等, 2018)。叶形的变化也影响到植物生物量的分配策略,叶面积减小,降低植物对碳和能源的消耗,大部分同化产物被运输到根部,促进根系生长(Lincoln Taizetal., 2015),这与本研究根茎比增加的结果相一致,根茎比增加利于植物在表层土壤水匮乏的干旱条件下扩大根系吸水面积(闫海霞等, 2011; 井大炜等, 2014)。
3.2 高温和干旱胁迫对杨树叶肉细胞超微结构的影响 从叶形态角度去讨论植物对环境的生长策略,只是根据叶片生长状态所做的推测,叶片形态性状的改变主要是由物种遗传特性所决定的(韩威等, 2014; 李东胜等, 2013)。在重度非生物环境因子胁迫下,植物的生长变化也有可能是生理过程受到干扰的表现。因此,通过电镜图像分析比较不同程度的胁迫对植物超微结构的损伤,可以更准确地区分逆境条件下植物的适应与受迫状态。干旱胁迫下植物细胞脱水,液泡变小,膨压降低。膨压驱动细胞伸展的过程对水分亏缺十分敏感,即使降幅很小,也可导致叶片生长缓慢,甚至完全停止生长。这与观察到的重度干旱胁迫下叶肉细胞变窄变小的趋势一致。同时观察到水分亏缺时叶绿体数量减少,说明干旱胁迫下植物生长受抑可能是光合细胞器减少和膨压共同作用的结果。与干旱相比,高温对细胞膜结构的损伤更严重,在干旱胁迫下相对稳定的叶绿体被膜在重度高温时有消解的趋势。在中度干旱胁迫下提高处理温度,细胞发生严重的内膜紊乱和胞质外渗现象。内膜系统的稳定是维持胞内正常生理过程的必要条件。高温可导致细胞膜损伤和酶失活,并通过改变叶绿体的膜特性和能量转移机制的解偶联影响光合速率(Lincoln Taizetal., 2015)。类囊体膜上的光系统Ⅱ对温度极其敏感,是高温抑制光合作用的关键环节(郝召君等, 2017)。对类囊体垛叠与松散及其功能发挥的研究指出,H+的传递受2个光系统、电子递体间距离及偶联程度的限制(钱露萍等,1985),片层的扩张会影响光反应的电子传递进而影响光合作用。
‘南林895杨’幼苗胁迫条件出现叶绿体内嗜锇颗粒增多和淀粉粒减少现象,这在不同物种也有类似情况(陈健辉等, 2011; Lianopoulouetal., 2014)。嗜锇颗粒作为一些特殊脂类分子合成与贮存的场所,与类囊体相偶联,在胁迫时脂类分子可作为电子载体保护类囊体使其免受自由基伤害(Grigorovaetal., 2012)。也有研究认为,嗜锇颗粒是类囊体降解膜脂聚集的产物,反映了胁迫条件下叶绿体受害程度加剧(万里强等, 2009; 杜克兵等, 2010; 周艳威等, 2018),本研究随胁迫程度增加,‘南林895杨’幼苗叶片嗜锇颗粒明显增多。逆境条件下植物的应激反应和代谢能力下降都会造成淀粉粒积累,而光合能力的降低会减少淀粉粒的数量(唐琪等, 2018; 汪贵斌等, 2008; 熊贤荣等, 2017; 郁慧等, 2011)。复合胁迫下细胞发生不同程度的皱缩,质壁分离严重。细胞内膜系统在重度胁迫下模糊甚至消失,胞质外渗,对胞内正常生理过程产生严重影响。在单一胁迫下较为稳定的细胞核核仁消失,染色质凝集。也有研究指出,复合胁迫与单一胁迫是2种完全不同的过程(Grigorovaetal., 2012),它对植物的影响也不是单一胁迫的简单叠加。干旱条件下植物通过蒸腾减少高温伤害的抗逆行为受到限制,单一胁迫下植物的适应机制在复合胁迫下可能失去原有效用。复合胁迫下‘南林895杨’生物量骤减,与超微结构受迫程度一致,高温、干旱的复合胁迫对植物的影响比单一胁迫更为严重。
4 结论
高温干旱胁迫下的叶片超微结构的损伤影响了‘南林895杨’扦插苗的光合作用和细胞内环境的稳定,正常的生理过程受到影响,并最终反映在生长形态的变化上。总体而言,无论是生理生长还是细胞结构层面的结果均表明,复合胁迫对扦插苗的影响要大于单一胁迫。在本研究中,超微结构的改变与扦插苗的生长指标表现出一致性,说明从形态和细胞学角度来揭示植物对于胁迫的响应机制是可行的。结合对生理生化指标,如抗氧化酶、光合参数的定量测定,可以更好地判断植物在不同程度胁迫下的适应与受迫情况。
