昆虫气味受体研究技术及其在林业昆虫中的应用研究进展*
2020-06-15申思凡孔祥波张苏芳
申思凡 张 真 孔祥波 刘 福 张苏芳
(中国林业科学研究院森林生态环境与保护研究所 国家林业和草原局森林保护学重点实验室 北京100091)
嗅觉在昆虫的生存和种群繁衍过程中发挥着重要的作用,昆虫通过嗅觉感受器识别周围环境中的气味分子,并将这些化学信号转变为神经电信号,从而调节自身的生物学过程。随着昆虫行为学、生物化学、分子生物学和昆虫电生理技术的发展,昆虫的嗅觉反应机制研究不断深入。研究发现,气味结合蛋白(odorant binding protein, OBP)、气味受体(odorant receptor, OR)、气味降解酶(odorant degrading enzyme, ODE)、化学感受蛋白(chemosensory protein, CSP)、感觉神经元膜蛋白(sensory neuron membrane protein, SNMP)等多种蛋白质都参与了气味识别过程,其中昆虫嗅觉感受的第一个步骤由存在于化感器腔内的气味结合蛋白完成,它与气味分子结合,并将其运送至嗅觉神经末梢(Vogtetal., 1999),而气味受体介导的气味分子与嗅觉感受器内的神经元的专一性结合是嗅觉识别的重要基础(Vosshalletal., 2000; Benton, 2006)。因此,气味受体研究是阐明昆虫嗅觉识别机制的重要部分,吸引众多昆虫学家不断探索,目前已经取得了较为深入的研究结果。前人已对气味受体(包括普通气味受体、信息素气味受体和非典型气味受体)的功能,与G蛋白偶联受体(G protein coupled receptor, GPCR)的联系,分子进化及信号转导机制等做了总结(乔奇等, 2008; 张春香等, 2011; 俞明明等, 2011; 尹淑艳等, 2013; Zhangetal., 2014)。笔者基于昆虫气味受体研究的已有结果,从其发现、特征、基因功能研究方法几个方面逐层总结了昆虫气味受体的功能和分子机制; 并着重分析了林业昆虫气味嗅觉识别和气味受体研究的进展和不足,以期为林业昆虫嗅觉识别机制研究提供参考。
1 OR基因的发现历史
1991年气味受体家族在大鼠(Rattusnorvegicus)嗅觉上皮细胞被发现(Bucketal., 1991); 1992年人类气味受体也被找到(Parmentieretal., 1992); 随后斑点叉尾鮰(Lctaluruspunctatus)等脊椎动物的气味受体基因通过同源克隆的方式得到(Ngaietal., 1993); 1995年,通过全基因组测序的方法筛选到秀丽隐杆线虫(Caenorhabditiselegans)的气味受体家族(Troemeletal., 1995)。之后,Clyne(1999)首次在果蝇中鉴定到第一个昆虫气味受体家族。后人不断对果蝇气味受体进行研究,其家族也逐渐扩大,共鉴定到62个受体基因(Vosshalletal., 2000; Jonesetal., 2005)。之后,冈比亚按蚊(Anophelesgambiae)(Hilletal., 2002)、家蚕(Bombyxmori)(Sakuraietal., 2004; Wanneretal., 2007)、意大利蜜蜂(Apismellifera)(Robertsonetal., 2006)、埃及伊蚊(Aedesaegypti)(Bohbotetal., 2007)全基因组测序成功,分别鉴定得到79、48、170、131个OR基因。
2 OR基因的结构和分类
昆虫气味受体是位于嗅觉感受神经元树突上的膜蛋白,由一个基因家族所编码,与脊椎动物或线虫的气味受体的基因序列同源性较低。目前,果蝇OR基因研究最为透彻,虽然对鳞翅目家蚕(Wanneretal., 2007)、烟芽夜蛾(Heliothisvirescens)(Soquesetal., 2010),双翅目冈比亚按蚊(Meijerinketal., 2001)、果蝇(Kielyetal., 2007)、瓜实蝇(Bactroceracucurbitae)(申建梅等, 2011)等,膜翅目意大利蜜蜂(Wanneretal., 2007)等昆虫也进行了一些研究,但研究的目标昆虫尚集中在一些具有重要经济或医学意义的昆虫种类上,如家蚕和烟芽夜蛾等。
昆虫OR由300~500个氨基酸组成,N端没有信号肽序列; 昆虫气味受体的结构与具有7个跨膜区域的脊椎动物的GPCR类似,但是不同于GPCR的膜拓扑结构,其N端位于细胞内且C端位于细胞外(Bentonetal., 2006)。昆虫的OR基因可以分为2类,一类是编码功能气味受体的基因,这类基因在不同昆虫间的同源性较低,另一类基因在不同昆虫间较为保守(Larssonetal., 2004),与传统的气味受体不同,这类基因不感受气味分子,而是作为一种共表达受体基因,与传统气味受体一起识别气味分子(Dobritsaetal., 2003)。这类受体最初在果蝇中被鉴定出来,被命名为Or83b (Vosshalletal., 2000),后来又在不同的昆虫中鉴定出了Or83b的同源受体,如冈比亚按蚊(Pittsetal., 2004)、埃及伊蚊(Meloetal., 2004)、烟芽夜蛾(Kriegeretal., 2003)等,当时被命名为不同的名字。为了方便交流,同时由于不同昆虫之间的这类基因同源性很高,所以目前已被统称为Orco(Odorant receptor co-receptor)(Vosshalletal., 2011)。
Orco是将传统OR定位于树突膜所必需的(Stengletal., 2013)。Orco与传统的气味受体通过保守的C末端区域相互作用形成OR-Orco复合物,这一新的配体门控离子通道,当Orco被基因打靶或RNA干扰沉默时,昆虫会表现出严重的嗅觉缺陷。阐明Orco在昆虫嗅觉识别中的作用机制可能为开发基于昆虫嗅觉行为干扰的新型害虫防治措施提供依据。Orco中的大多数氨基酸残基在昆虫序列中保守性很好。Jones等(2005)利用GAL4-UAL系统,将与果蝇Orco基因直系同源的地中海实蝇(Ceratitiscapitata)、冈比亚按蚊和美洲棉铃虫(Helicoverpazea)的Orco基因在突变果蝇品系的相应嗅觉感受神经元(olfactory sensory neuron, OSN)中表达,转基因果蝇可以恢复其相应的嗅觉功能。Mitsuno等(2008)也得出了相应的结论,他们在对小菜蛾(Plutellaxylostella)、黏虫(Mythimnaseparata)和瓜绢螟(Diaphaniaindica)相应的OR进行功能测试的时候,将与之共注射的自身表达的Orco基因换成了家蚕的Orco基因,发现其对气味分子的感受谱不变,显示出了Orco基因的高度保守性。
3 OR基因功能研究主要方法
目前,对OR基因的研究,主要集中于鉴定表达及功能方面。关于鉴定,常用的方法有原位杂交(Grosse-Wildeetal., 2010)、RT-PCR(Mitsunoetal., 2008)、转录组测序(Zhangetal., 2014)、甚至基因组测序(Mckennaetal., 2016); 表达量研究一般通过实时定量PCR(quantitative real time polymerase chain reaction, Q-PCR)或者转录组测序检测各个气味受体基因在各个组织中的表达量(Patchetal., 2009; Zhangetal., 2017)。由于OR属于膜蛋白,其功能研究主要依赖于几种膜蛋白表达系统,如HEK293细胞(Grosse-Wildeetal., 2007)、 爪蟾卵母细胞体外表达系统(Mitsunoetal., 2008)、 果蝇空神经元系统(Montagnéetal., 2012)等。同时,RNAi (RNA interference)与Crispr/Cas9 (clustered regularly interspaced short palindromic repeats/CRISPR-associated sequence)系统也逐渐在OR功能研究中得到应用(Zhuetal., 2013; Lietal., 2016)。
3.1 爪蟾卵母细胞体外表达系统
非洲爪蟾(Xenopuslaevis)的卵母细胞是应用最早的基因功能表达体系之一。Gurdon等(1981)首次将纯化的DNA注射到爪蟾卵母细胞获得了正确的转录和表达,随着分子生物学的发展,其应用又有进一步的扩展。Mitsuno等(2008)将小菜蛾、黏虫、瓜绢螟OR1和其Orco的cRNA共同注射到爪蟾卵母细胞中,成功表达,随后利用双电极电压钳实验检测了3个OR对7种气味分子的反应,小菜蛾对Z11-16:Ald的反应最强,黏虫对Z11-16:Ac的反应最强,瓜绢螟对E11-16:Ald的反应最强。该技术在昆虫OR功能研究中得到了广泛的应用,完成了多种昆虫OR功能鉴定,如玉米螟(Ostrinianubilalis)(Wanneretal., 2010)、棉铃虫(Helicoverpaarmigera)和烟青虫(Heliothisassulta)(Jiangetal., 2014; Caoetal., 2016)、甜菜夜蛾(Spodopteraexigua)(Liuetal., 2013; Liuetal., 2014)、斜纹夜蛾(Spodopteralitura)(Zhangetal., 2013; 2015)等。
3.2 其他异源细胞表达系统
HEK293细胞是一个很常用的研究外源基因功能的细胞株。Grosse-Wilde等(2007)将烟芽夜蛾的OR13受体在HEK293细胞中成功表达,并通过钙离子成像的方法检测到了气味分子刺激时细胞内钙离子浓度的变化; 发现其对主要的性信息素组分Z11-16:Ald反应最强,对其他几种气味分子也可以产生反应,推测OR13可能为性信息素受体。
Sf9细胞是草地贪夜蛾(Spodopterafrugiperda)细胞,是一种昆虫表达系统的宿主细胞。Jordan等(2009)鉴定了苹淡褐卷叶蛾(Epiphyaspostvittana)的3个OR基因,钙离子成像结果显示EpOR1和EpOR3都能识别植物产生的一系列萜类和苯甲酸酯,EpOR1对水杨酸甲酯最敏感,这是花卉气味的常见成分,也是植物受到昆虫和病原体侵袭时产生的重要信号化合物,EpOR3能很好地识别单萜柠檬醛,并引发驱避雌性飞蛾排卵的活性; 而EpOR2与黑腹果蝇的共同受体Orco是直系同源的。
3.3 果蝇空神经元系统
果蝇是研究复杂生物过程中遗传和细胞生物学通路的一个重要模型,用分子遗传学方法构建UAS/GAL4系统可以将任何基因在果蝇的特定组织和细胞内进行表达。Montagné等(2012)将灰翅夜蛾(Spodopteralittoralis)的性信息素受体OR6成功在果蝇体内表达,并通过单感记录检测其对气味分子产生的反应,发现其对Z9, E12-14:OAc反应最强。之后,de Fouchier等(2017)首次系统分析了鳞翅目灰翅夜蛾的全部OR基因,将47个候选OR基因成功表达30个,其中的17个OR基因成功找到了气味配体。每个OR都存在于相应的OSN中,对表达相应气味受体的OSN进行功能分析,结果显示其对气味的识别与OR基因对气味的识别一致,表明果蝇空神经元系统的可信度很高。随后对已验证功能的OR基因进行了系统发育分析,最基础的谱系表现了低的平均遗传距离,它们对芳香烃最敏感,其次是萜烯类受体,而脂肪族受体属于具有最高平均遗传距离的OR谱系。这表明芳香族化合物的受体首先出现并且在鳞翅目进化过程中更加保守,而萜烯和脂肪族化合物的受体最近出现并且进化得更快(特别是脂肪族受体,包括信息素受体)。其中,SlitOR36是一个例外,该受体对所有活性配体均表现出高响应阈值,表明其关键配体仍有待确定。
3.4 RNAi与Crispr/Cas9系统在OR中的应用
RNAi是一种转录后基因沉默机制,由双链RNA(double-stranded RNA, dsRNA)进入细胞形成的。Zhu等(2013)克隆了热带家蚊(Culexquinquefasciatus)2个气味受体基因,OR37和OR99,分别对4-甲基苯酚和4-乙基苯酚有反应,实验证明产卵期的雌蚊在含有这2种信息素的容器里的产卵量明显高于只有水的容器; 将CquiOR37/99-dsRNA,β-galactosidase-dsRNA或水分别注射到蛹的血淋巴中,Q-PCR分析显示个体差异,但CquiOR37/99-dsRNA处理的家蚊中CquiOR37和CquiOR99转录物水平显着降低,注射水的雌蚊和用对照基因处理的雌蚊在含有4-乙基苯酚的盘中比在水盘中产生更多的卵,而CquiOR37/99-dsRNA处理的雌蚊产生正常数量的卵,但不能区别于对照组; 该研究首次将4-乙基苯酚的特异性气味受体与热带家蚊的产卵增加联系起来。
基因组编辑是基因功能分析的重要工具,并表现出了在害虫防治方面的潜在应用。与锌指核酸酶(zinc-finger nuclease, ZFN)和类转录激活因子效应物核酸酶(transcription-activator-like effector nuclease, TALEN)相比,Crispr/Cas系统能够更精确,更高效地进行基因组编辑,并且更加便于实施。基于Crispr/Cas9系统的昆虫功能基因组学研究为害虫防治提供了新思路,不仅应用于模式昆虫(如飞蝗、家蚕等),在非模式昆虫[如苹果蠹蛾(Cydiapomonella)等]中也得到了成功应用。Li等(2016)使用Ⅱ型Crispr/Cas系统成功设计gRNA (guide RNA)的靶序列破坏编码气味受体共受体(Orco)的基因并检查气味受体途径在东亚飞蝗(Locustamigratoria)中的作用,为蝗虫功能遗传学研究和害虫管理产生功能丧失突变体提供了一种简单而有效的方法。Liu等(2017)敲除了家蚕的Orco基因,获得了纯合突变体,单感记录显示这种纯合突变体的触角对家蚕的2种性信息素蚕蛾醇和蚕蛾醛均没有反应。成虫的交配试验结果显示,Orco突变体无法响应野生雌蛾释放的性信息素,交配行为明显受损; 幼虫的取食试验结果显示,Orco突变体表现出对桑叶的选择缺陷。综上,Orco基因的敲除严重破坏了家蚕的嗅觉系统,这项研究为昆虫嗅觉系统提供了新见解,也为农林复合害虫防治提供了范例。Garczynski等(2017)发现苹果蠹蛾的OR1基因被靶向敲除后,突变雌性的繁殖力受到了严重的影响。
4 林业类害虫的嗅觉识别分子机制
嗅觉感受的分子机制研究将为农林害虫防治提供新的线索,目前在重要农林害虫中的OR鉴定和功能研究情况如表1所示。可以发现,林业害虫嗅觉识别机制和OR的研究还处于起步阶段,大部分还限于OR等嗅觉基因的鉴定,功能分析等深入研究则较少。以下是几种较为重要的森林害虫嗅觉机制的研究情况。
4.1 鞘翅目害虫
白蜡窄吉丁(Agrilusplanipennis)在东北亚(中国、日本、朝鲜、蒙古)、美国、加拿大及俄罗斯远东地区等均有分布,在国内分布于黑龙江、吉林、辽宁、河北、北京、天津、台湾等地(杨忠岐等, 2018),严重危害我国北方重要的用材树种水曲柳(Fraxinusmandshurica)和园林绿化树种绒毛白蜡(Fraxinusvelutina)。2013年,Mamidala等(2013)首次对白蜡窄吉丁的触角进行了转录组测序,得到了9个OBP基因、2个OR基因、1个SNMP基因和134种气味/异生素降解酶,包括细胞色素P450、谷胱甘肽-S转移酶、酯酶等; 并通过Q-PCR揭示了这些基因在不同发育阶段雌雄体内的发育和性别偏向的表达模式。
Mitchell等(2012)对胡桃胭脂天牛(Meyacyllenecaryae)的转录组分析得到了57个0R基因,并利用爪蟾卵母细胞表达系统对其中的3个OR基因进行了功能测定,其中OR3对(S)-2-甲基-1-丁醇敏感,OR5对2-苯乙醇敏感,OR20对(2S,3R)-2,3-己二醇敏感。
光肩星天牛(Anoplophoraglabripennis)是危害树木生长的蛀干害虫,在我国陕西、宁夏、内蒙古、河南、山东、湖南、湖北、辽宁、吉林等地发生十分严重,近年来在新疆、西藏等西部偏远地方也已被发现,每年都造成大量的木材损失,是一种重要的林业害虫(杨忠岐等, 2018)。2016年,McKenna等(2016)完成了光肩星天牛的基因组测序与注释工作,并进行了基因表达分析,Hu等(2016)也对光肩星天牛进行了转录组分析,鉴定得到42个OBP基因、12个CSP基因、14个信息素降解酶(pheromone degrading enzyme, PDE)基因、1个ODE基因、37个OR基因、11个味觉受体(gustatory receptor, GR)基因、2个SNMP基因和4个离子型受体(ionotropic receptor, IR)基因,并通过CSP基因和PBP基因在触角中的正确表达,证明了转录组数据的真实性。Mitchell等(2017)在此基础上,鉴定得到了120个OR基因、1个Orco基因和11个可能的OR基因。昆虫行使嗅觉功能时,表达OR基因的嗅觉感受神经元(OSN)通过其轴突连接大脑触角叶,相同OSN类型的轴突汇聚在同一个嗅觉小球中,将信号传递给大脑。OSN轴突的数量多少决定了嗅觉小球的大小,这同时又可以反映出昆虫接受气味信息的多少。这项研究将基因组学与神经解剖学联系起来,对气味受体及其配体从一个新的角度进行了诠释。
云杉八齿小蠹(Ipstypographus)和中欧山松大小蠹(Dendroctonusponderosae)均为重要的针叶树害虫,通过聚集信息素来调节对宿主的入侵行为,在此过程中嗅觉起着重要的作用。Andersson等(2013)对这2种小蠹虫的转录组进行了分析,在云杉八齿小蠹中找到了15个OBP基因、6个CSP基因、3个SNMP基因、43个OR基因、6个GR基因和7个IR基因,在中欧山松大小蠹中找到了31个OBP基因、11个CSP基因、3个SNMP基因、49个OR基因、2个GR基因和15个IR基因。对这几种蛋白质的系统发育分析表明,只有OR蛋白具有明显的谱系特异性,这表明可能OR与感官特异性、特异生态位的适应和化学信息素的共享等联系更紧密。
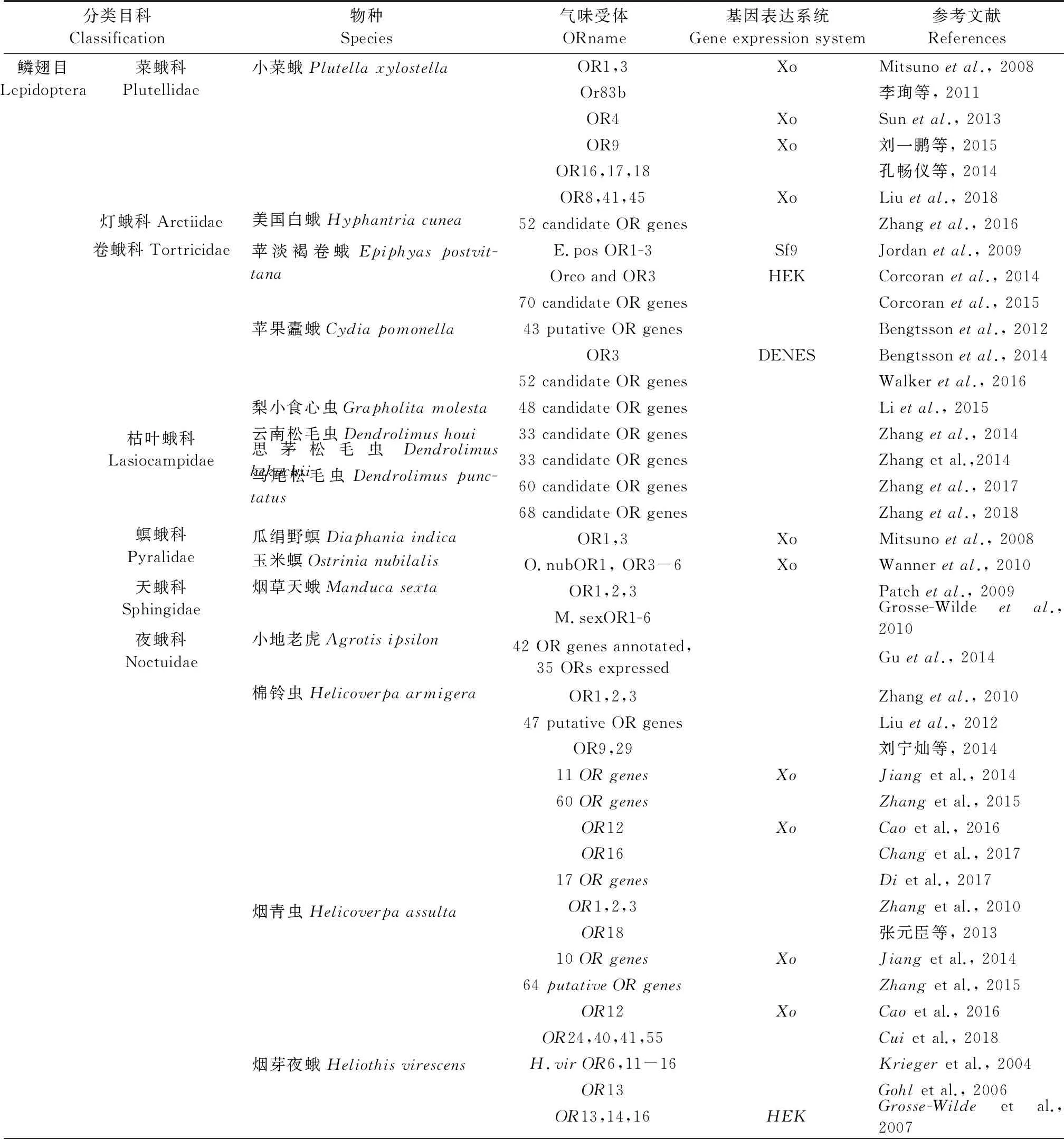
表1 农林类害虫气味受体鉴定和功能研究情况
续表1 Continued
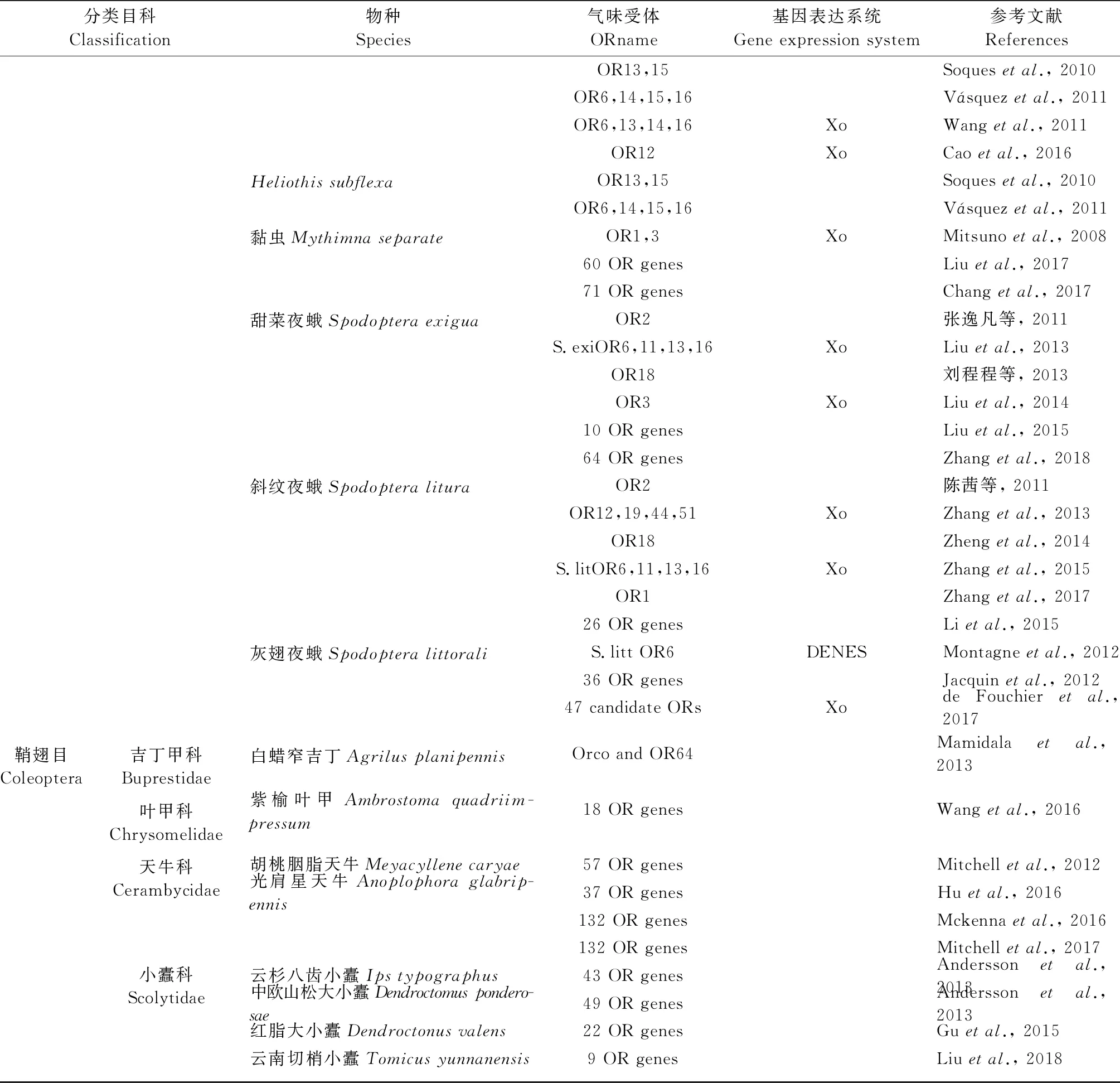
分类目科Classification物种Species气味受体ORname基因表达系统Gene expression system参考文献ReferencesHeliothis subflexa黏虫Mythimna separate甜菜夜蛾Spodoptera exigua斜纹夜蛾Spodoptera litura灰翅夜蛾Spodoptera littoraliOR13,15Soques et al., 2010OR6,14,15,16Vásquez et al., 2011OR6,13,14,16XoWang et al., 2011OR12XoCao et al., 2016OR13,15Soques et al., 2010OR6,14,15,16Vásquez et al., 2011OR1,3XoMitsuno et al., 200860 OR genesLiu et al., 201771 OR genesChang et al., 2017OR2张逸凡等, 2011S.exiOR6,11,13,16XoLiu et al., 2013OR18刘程程等, 2013OR3XoLiu et al., 201410 OR genesLiu et al., 201564 OR genesZhang et al., 2018OR2陈茜等, 2011OR12,19,44,51XoZhang et al., 2013OR18Zheng et al., 2014S.litOR6,11,13,16XoZhang et al., 2015OR1Zhang et al., 201726 OR genesLi et al., 2015S.litt OR6DENESMontagne et al., 201236 OR genesJacquin et al., 201247 candidate ORsXode Fouchier et al., 2017鞘翅目Coleoptera吉丁甲科Buprestidae叶甲科Chrysomelidae天牛科Cerambycidae小蠹科Scolytidae白蜡窄吉丁Agrilus planipennis紫榆叶甲Ambrostoma quadriim-pressum胡桃胭脂天牛Meyacyllene caryae光肩星天牛Anoplophora glabrip-ennis云杉八齿小蠹Ips typographus中欧山松大小蠹Dendroctomus pondero-sae红脂大小蠹Dendroctonus valens云南切梢小蠹Tomicus yunnanensisOrco and OR64Mamidala et al., 201318 OR genesWang et al., 201657 OR genesMitchell et al., 201237 OR genesHu et al., 2016132 OR genesMckenna et al., 2016132 OR genesMitchell et al., 201743 OR genesAndersson et al., 201349 OR genesAndersson et al., 201322 OR genesGu et al., 20159 OR genesLiu et al., 2018
①Xo:爪蟾卵母细胞Xenopus oocyte; HEK:HEK293细胞HEK293 cell; DENES:果蝇空神经元系统Drosophila empty neuron expression system.
红脂大小蠹(Dendroctonusvalens)对油松(Pinustabulaeformis)造成严重的危害,原产北美; Gu等(2015)对红脂大小蠹的转录组分析得到了可能的21个OBP基因、6个CSP基因、4个SNMP基因、22个OR基因、4个GR基因和3个IR基因。小蠹科中,在红脂大小蠹中鉴定得到的OR基因比云杉八齿小蠹和中欧山松大小蠹中都少,但是系统发育分析结果显示,红脂大小蠹的嗅觉基因大部分与中欧山松大小蠹的嗅觉基因聚类。在北美,红脂大小蠹和中欧山松大小蠹的生境相似,共享相似的宿主挥发物、信息素挥发物以及非宿主挥发物,这可能解释在系统发育分析中2种小蠹大部分聚类的现象。
云南切梢小蠹(Tomicusyunnanensis)分布于我国西南部,对松林造成了严重的危害。Liu等(2018)对云南切梢小蠹的3个发育阶段(幼虫、蛹、成虫)的化学感受基因做了差异表达分析,并找到了45个OBP基因、12个CSP基因、9个OR基因、8个GR基因和3个IR基因。
紫榆叶甲(Ambrostomaquadriimpressum)亦是一种主要的林木害虫,目前尚未发现有效且环境友好的化学防治方法来控制这种害虫。Wang等(2016)对紫榆叶甲腿部及触角进行了转录组分析,经生物信息学分析,同源分析及后续RT-PCR和Q-PCR等试验,最终得到了15个OBP基因、9个CSP基因,18个OR基因、6个IR基因和2个SNMP基因。其中,AquaOBP1/2/4/7/C1/C6、AquaCSP3/9、AquaOR8/9/10/14/15/18/20/26/29/33、AquaIR8a/13/25a显示嗅觉特异性表达,这些基因可能在紫榆叶甲的嗅觉相关行为中发挥关键作用;AquaOBP4/C5、AquaOBP4/C5、AquaCSP7/9/10、AquaOR17/24/32和AquaIR4在雄性触角中表达量较高且具有特异性,表明这些基因与性别特异性行为有关。
4.2 鳞翅目害虫
云南松毛虫(Dendrolimushoui)和思茅松毛虫(Dendrolimuskikuchii)是2个及其相近的物种,对中国西南地区的针叶林危害严重,尽管鳞翅目物种的嗅觉研究工作较多,但是关于2个姐妹种的嗅觉识别机制还研究的很少。Zhang等(2014)对这2大食叶害虫做了转录组分析,同源比较在云南松毛虫中得到了23个OBP基因、17个CSP基因、2个SNMP基因、33个OR基因和10个IR基因,在思茅松毛虫中得到了27个OBP基因、17个CSP基因、2个SNMP基因、33个OR基因和9个IR基因,对未来的功能研究提供了依据。马尾松毛虫(Dendrolimuspunctatus)也是一种危害严重的食叶害虫,其生理生态研究较多,但有关其发育的分子机制还了解的不多。Yang等(2016)对马尾松毛虫进行了转录组分析,构建了卵、幼虫、蛹、成虫4个阶段的基因表达谱。Zhang等(2017)用新一代测序技术对马尾松毛虫不同发育阶段和不同器官进行了转录组分析,共得到171个可能的化学感受基因,包括53个OBP基因、26个CSP基因、2个SNMP基因、60个OR基因、18个IR基因和12个GR基因,其中触角中的嗅觉基因表达含量最高,其他组织中等表达,而脂肪体、中肠、卵巢和精巢组织中也发现了一定量的嗅觉基因的表达(Zhangetal., 2017)。随后,Zhang等(2018)进一步对马尾松毛虫羽化、求偶、交配、交配后4个交配阶段进行了转录组分析,得到了相关化学感受基因在其交配期间3种不同的表达模式,通过后续的系统发育分析、基序表达模式分析等推测了相关化学感受基因的功能; 且在之前研究中的6个OBP基因、1个CSP基因、23个OR基因、1个GR基因和8个IR基因获得了全长序列,8个OR基因、9个GR基因和5个IR基因被新鉴定。
美国白蛾(Hyphantriacunea)繁殖率高,传播范围广,对我国的森林甚至农作物造成了严重的危害,但关于其寄主定位及交配选择嗅觉机制的分子基础知之甚少。Zhang等(2016)对美国白蛾进行了转录组分析,得到了30个OBP基因、17个CSP基因、52个OR基因、14个IR基因、9个GR基因和2个SNMP基因,并通过RT-PCR和Q-PCR系统分析了OBP和CSP的表达模式,为更深入研究嗅觉机制提供了新的线索。
5 结语
有关昆虫嗅觉分子机制的研究在近年来有新进展,气味受体作为昆虫行使嗅觉功能的一种重要的蛋白质,是昆虫嗅觉研究的重点部分。从昆虫异源细胞表达系统、爪蟾卵母细胞表达系统、果蝇空神经元表达系统、RNAi技术,到Crispr/cas9等新型技术在气味受体功能研究中的应用,使人们对气味受体功能的认识逐渐深入。除了对一些模式昆虫,如家蚕、黑腹果蝇等的深入研究外,一些农业类重要害虫的研究也获得了较为广泛的关注。
林业类害虫嗅觉识别分子机制的研究在近几年逐渐展开,目前主要集中在基因鉴定方面,功能研究和应用开发涉及较少。未来展开对林业害虫气味受体功能的研究,可以帮助人们从分子角度深入了解昆虫的嗅觉识别机制,有望以此为基础开发效率更高的引诱剂,推出林业害虫防治新技术。
