基于亲本对条锈病敏感性预测小麦杂交种的抗性
2020-06-03周天宇李姜玲杨澜阮仁武杨宇衡李中安
周天宇,李姜玲,杨澜,阮仁武,杨宇衡,李中安
基于亲本对条锈病敏感性预测小麦杂交种的抗性
周天宇1,李姜玲1,杨澜1,阮仁武2,杨宇衡3,李中安1
(1西南大学柑桔研究所,重庆 400712;2西南大学农业与生物技术学院,重庆 400716;3西南大学植物保护学院,重庆 400716)
【】通过亲本条锈病的抗性评价预测F1代杂交种的抗病性,增强杂交小麦抗病育种的可预见性。以CYR23、CYR31、CYR33、CYR34 4个小麦条锈菌(f. sp.)生理小种作为供试菌源,感病小麦品种铭贤169作为阴性对照,通过成株期混合接种,对13份恢复系(父本)材料和21份不育系(母本)材料及其F1代杂交种进行抗病性鉴定,并利用、、、、、、等抗条锈基因的分子标记或基因标记对其可能携带的抗条锈基因进行分子检测。同时通过半定量PCR方法在亲本及部分F1代植株的成株期进行条锈菌侵染量测定。所有材料均未鉴定到、、,多存在于四川品系,、多存在于北方品系,本研究所有恢复系材料均未鉴定到。亲本抗条锈基因在F1代杂交种得到了聚合,符合遗传规律,表明分子标记可以用于杂交小麦抗病辅助选育。来自四川的恢复系及其F1代杂交种整体表现优良抗性,推测其具有纯合显性的抗条锈基因,同时,这些小麦材料可以用于我国小麦抗条锈育种。F1代的实际鉴定反应型趋于亲本反应型的平均值,二元回归分析结果表明亲本和F1代的反应型之间有显著相关性(2=0.812)。来自四川的恢复系及其F1代对新毒性小种CYR34表现优良抗性,但亲本中未检测到、,推测其可能含有未知的抗条锈病基因。利用半定量PCR方法对所有恢复系、不育系和部分F1代杂交种分别进行了供试条锈菌生理小种的菌量测定,结果显示所有亲本及其杂种中均未检测到CYR23,恢复系15CA50、不育系17L6078和15L7128有少量CYR31侵染,恢复系川13品6、MR1101和川麦98及其F1代杂交种未检测到CYR33、CYR34。同时发现,对不同生理小种抗性互补的亲本能有效提高F1代的抗病性。根据双亲对条锈病的反应型可以预测其F1代杂交种的抗性水平,双亲的抗病水平越高,其F1代杂交种的抗性就越好。同时可以选用具有不同条锈病生理小种抗性互补的亲本来提高F1代杂交种的抗性水平。研究结果有助于探究亲本与F1代杂交种之间的抗病规律,同时为杂交小麦抗病育种提供可参考的实践方案。
杂交小麦;条锈病;抗性预测;小麦条锈菌;生理小种
0 引言
【研究意义】小麦是全球第二大粮食作物,随着世界人口不断增加而耕地面积逐年减少,预计到2050年全球粮食需求将增长一倍,因此粮食安全非常重要[1-4]。小麦条锈病严重威胁小麦安全生产,因此筛选小麦抗条锈育种材料并预测杂交小麦的抗锈性,对我国杂交小麦抗病育种以及粮食安全具有重要意义。【前人研究进展】20世纪60年代发现小麦雄性不育系和恢复系,这给小麦的杂种优势利用带来了希望[5]。近年来,杂交小麦迅速发展,强优势组合在生产上开始大面积示范[6]。杂种优势是指杂合体在一种或多种性状上优于两个亲本的现象,在自然界广泛存在,其表现形式为高产、抗病、较高的肥料利用率、根系渗透性、种子结实率,以及对非生物胁迫的强忍耐性等[7-8]。利用杂种优势一直被认为是提高粮食产量的重要途径,在水稻、玉米、高粱等作物上已取得显著效果[9-10]。70年来中国水稻品种实现了矮杆化、杂交化、优质化的3次跨越,完成6次新品种更新换代[11]。小麦的杂交化经历了漫长的过程,但仍未取得质的突破,还处于研究的初始阶段。小麦杂交化除了提高小麦单位面积产量外,降低生产成本已成为一个重要的育种目标,以小麦基因组为基础,利用分子标记辅助育种实现了部分农艺性状的早期选择,可以更好地培育生产上需要的新品种[12]。此外,通过亲本和F1代多个性状指标(如粒长、粒重等)建立线性回归关系对F1代进行优势预测已经在杂交水稻上得以证明[13]。小麦抗病育种对于小麦稳产、增产具有重要意义,农作物抗病基因的分离鉴定直接用于品种改良和病害绿色防控[14]。关于杂交小麦抗叶锈病、条锈病、黑穗病及白粉病的规律研究已有相关报道[15]。杂交小麦F1代抗白粉病研究表明,杂种一代与亲本的抗性表现基本一致,符合遗传规律[16]。将感小麦叶锈病的父本材料分别与不同反应型的母本材料杂交发现,抗叶锈母本的杂交后代大多数都表现抗病,并且这些杂合体有很强的杂种优势[17]。通过对单杂交种群体抗赤霉病分析,发现亲本对赤霉病表现的中亲值可以作为单杂交种对赤霉病表现的指标,并且亲本综合配合力与杂交种对赤霉病抗性有很高相关性[18]。Ali等[19]通过引入外源抗小麦条纹花叶病毒基因,并与其他基因组合提高了后代的抗性。利用分子标记技术将含有小麦抗白粉病基因(、)和抗小麦黄花叶病基因()的亲本进行杂交,构建F2代群体,在F3代群体中筛选出的23个双抗基因聚合体对白粉病免疫,对黄花叶病高抗,可用于双抗聚合品种的筛选或作为抗病中间亲本[20]。崔彩红等[21]利用分子标记技术定向培育了4个含有、4个含有、1个同时含有、的小麦新品系,这些品系农艺性状良好,可作为今后小麦抗秆锈育种的亲本。以上研究表明杂交小麦在抗病育种中能够发挥巨大作用,有助于加速小麦抗病育种的进程。小麦条锈病是由小麦条锈菌(f. sp.,)引起的、由气流传播的世界性重要病害,严重危害小麦产量[22]。该病原菌具有活体专性寄生、喜冷凉的特点,随降雨、露水等侵染小麦[23]。利用抗病品种是防治小麦条锈病最经济、最环保的方式,但小麦品种抗病基因单一化、病原菌生理小种变异强,再加上抗病品种选育慢,故造成小麦条锈病大流行[24],而且在杂交小麦的抗条锈病研究上未见系统报道。【本研究切入点】以小麦条锈病和小麦核不育杂交组合及双亲为研究对象,筛选含有不同的抗条锈病基因或对不同条锈病生理小种有抗性的亲本配制杂交组合,通过抗性评价明确杂交种抗性与双亲抗性间的关系。【拟解决的关键问题】通过分析亲本及F1代的反应型,鉴定抗条锈病基因及亲本、部分F1代成株期不同小种侵染量,预测杂交组合对条锈病的抗性,为杂交小麦亲本筛选和选育提供理论依据。
1 材料与方法
试验于2018—2019年在西南大学完成。
1.1 试验材料
供试小麦恢复系包括小偃22、陕987、高大1号、伟隆121、15CA50、丰德存5号、西农807、MR1101、MY13-3、川14品16、川13品6、川麦93、川麦98。不育系包括21份不育系,以及91份亲本杂交产生的F1代。条锈病感病对照品种为铭贤169,小偃22、高大1号、丰德存5号、西农807、伟隆121由陕西杨凌伟隆农业科技有限公司提供,MR1101、MY13-3由绵阳市农业科学院小麦研究所提供,川14品16、川13品6、川麦93、川麦98由四川省农业科学院作物研究所提供,15CA50由中国农业科学院棉花研究所提供。不育系及F1代由笔者实验室选育和配制,其中不育系为蓝标型核不育,不育基因为[25]。
供试菌种:CYR23、CYR31、CYR33、CYR34均在西南大学植物保护学院组培室利用感病品种铭贤169繁育获得。
1.2 方法
1.2.1 抗病基因分子鉴定 利用小麦抗条锈基因分子标记检测亲本中抗条锈基因的分布情况,所检测的抗病基因为、、、、、、,所用分子标记引物均由上海生工生物工程技术服务有限公司合成(表1[26-28])。采用CTAB法提取小麦基因组DNA[29],测定浓度后稀释至100 ng·μL-1备用。单个PCR检测体系总体积为20 μL,其中包含2×Taq Master Mix 10 μL,引物终浓度为2 mmol·L-1,DNA模板100 ng,ddH2O补足20 μL。产物均用2.0%的琼脂糖凝胶进行检测,并在凝胶成像仪上观察拍照。

表1 小麦抗条锈病基因的引物序列
Y15K1_F2/uhw301R为Kinase I引物,W_2F/W_2R为Kinase II引物Y15K1_F2/uhw301R are the primers ofKinase I, W_2F/W_2R are the primers ofKinase II
1.2.2 抗条锈病鉴定 每份小麦材料种植1行,行长1 m,行距0.25 m,每隔20个品种种植1行铭贤169作为感病对照,供试品种中间垂直种植1列感病品种铭贤169作为诱发行。分别称取0.1 g不同小麦条锈菌生理小种,混合配成1 L的孢子悬浮液,在小麦拔节期对诱发行进行混合接种。小麦进入抽穗期后,按0—9级标准记载反应型(infection type,IT),累计调查3次,以最高等级作为病情级数,根据IT抗病级别分为4个类型[30]:抗病(resistance,R,IT:0—3级)、中抗(moderate resistance,MR,IT:4—6级)、中感(moderate susceptibility,MS,IT:7级)、感病(susceptibility,S,IT:8—9级)。
1.2.3 亲本-F1代反应型的二元回归分析 以亲本成株期反应型为自变量,F1代成株期反应型为因变量,用SPSS软件对反应型之间的相关性进行二元回归分析。
1.2.4 亲本成株期小种侵染量鉴定 根据半定量PCR方法[31],利用小麦条锈菌生理小种特异分子标记引物鉴定不同生理小种对小麦亲本材料的侵染量。所鉴定的小麦条锈菌生理小种为CYR23、CYR31、CYR33、CYR34,所用分子标记引物均由上海生工生物工程技术服务有限公司合成。利用小麦条锈菌延伸因子特异引物EF作为参照[32](表2)。
小麦条锈菌DNA提取:根据成株期鉴定结果,采集发病的小麦叶片,准确称取叶片发病部位0.02 g,利用CTAB法提取总DNA,测定浓度后稀释至100 ng·μL-1备用。同时利用CTAB法提取条锈菌生理小种CYR23、CYR31、CYR33、CYR34的DNA作为阳性对照,测定浓度后稀释至100 ng·μL-1备用。
半定量PCR:将提取的发病叶片DNA用引物EF进行扩增,寻找不同材料扩增至相同亮度时的循环数,将每份材料按照确定的循环数分别用小种特异引物进行扩增,同时将纯小种模板进行所需循环数的扩增,单个PCR体系为2×Taq Master Mix 10 μL,引物终浓度2 mmol·L-1,DNA模板100 ng,ddH2O补足20 μL,重复3次。用软件Image J将条带转化为灰度值,计算每份材料中每个小种的侵染量。

表2 小麦条锈菌生理小种特异性引物序列
2 结果
2.1 亲本抗病基因分子检测
恢复系中不含、、,小偃22、陕987、高大1号含,陕987、高大1号、伟隆121、川麦98、MY13-3含,川14品16、川13品6、川麦93、川麦98、MR1101含(图1)。不育系中不含、、、抗病基因,在不育系中检测率比较高,占91%,17L9154、17L9160、17L9210、17L6065、17L6067含,17L6085、17L7030、17L7106、17L7123、17L7152含(图2)。来自四川的恢复系及其F1代表现出优良抗性,推测其中含有对CYR34未知的抗条锈基因。MY13-3及一些不育系未检测到但抗性优良,推测其含有未知的抗性基因(表3)。
2.2 亲本与F1代抗病表型
恢复系中反应型为抗病的包括川14品16、川13品6、川麦93、川麦98、MR1101、MY13-3;中抗的包括陕987、伟隆121;中感的包括高大1号、15CA50、丰德存5号、西农807;感病的为小偃22。不育系中反应型为抗病的包括15L7152、17L6062、17L6065、17L6067、17L7106、17L7123、17L7140、17L9160、17L9195、17L9210;中抗的包括08L5070、15L7109、15L7128、17L6019、17L6078、17L6085、17L7030、17L9066、17L9154、17L9163、17L9217(表3);F1代杂交种的反应型趋于亲本反应型的平均值(表4)。通过对亲本和F1代杂交种的反应型进行二元回归分析,得到2=0.812,表明F1代杂交种反应型和亲本有很大的相关性(图3)。综合以上结果,双亲都表现抗病,则杂交种也表现抗病;双亲均表现感病或抗性较差,其杂交种的抗性就差,但双亲互补的例外;双亲之一感病,则杂交种抗病表现介于两个亲本之间,绝大多数趋于中亲值。

表3 亲本抗病表现
“-”:未鉴定到本研究中的7个基因 The 7genes of this study are not detected

表4 F1代杂交种抗病表现
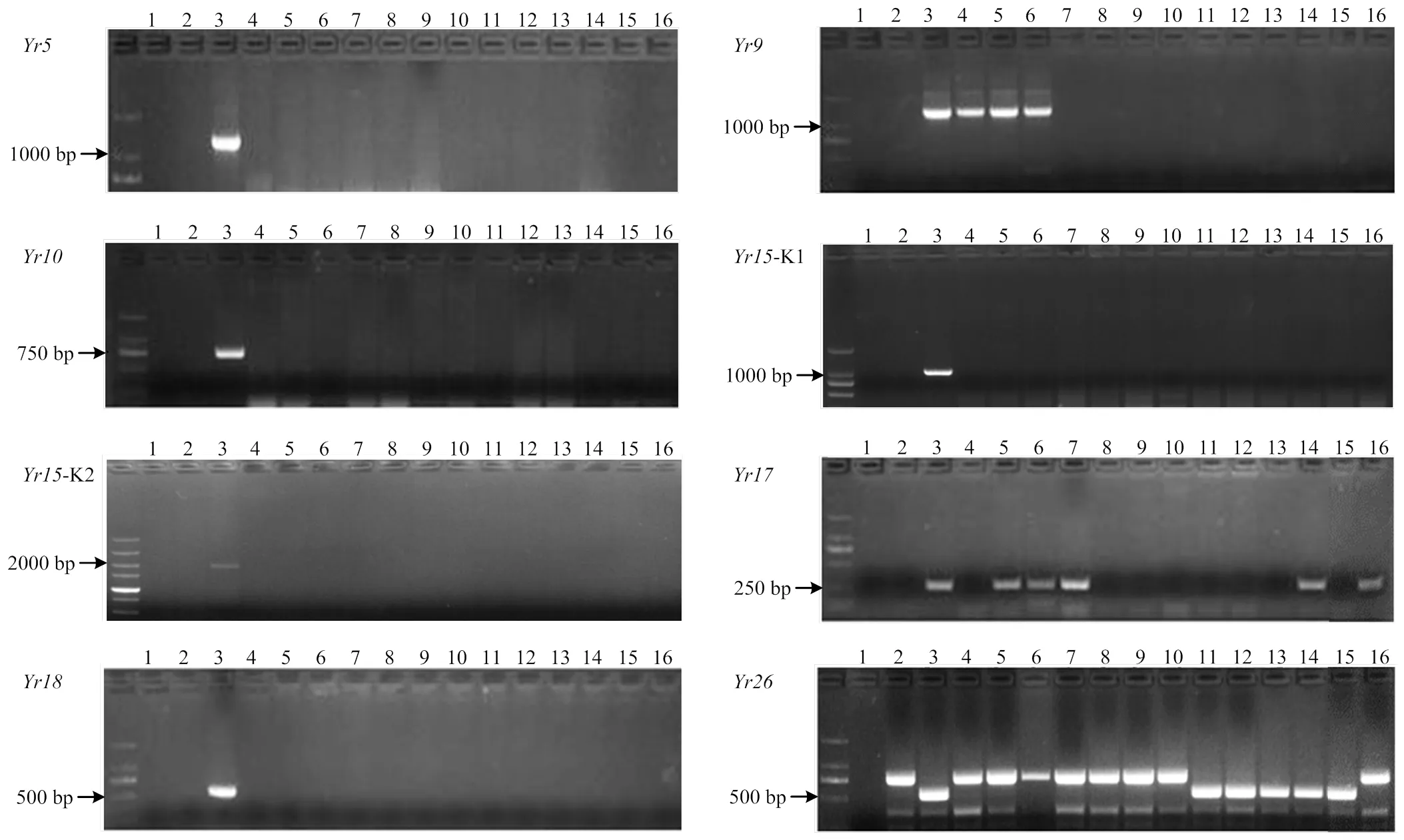
1:ddH2O;2:Avocet S;3:Yr;4:小偃22 Xiaoyan 22;5:陕987 Shaan 987;6:高大1号Gaoda No.1;7:伟隆121 Weilong 121;8:15CA50;9:丰德存5号 Fengdecun No.5;10:西农807 Xinong 807;11:川14品16 Chuan 14 pin 16;12:川13品6 Chuan 13 pin 6;13:川麦93 Chuanmai 93;14:川麦98 Chuanmai 98;15:MR1101;16:MY13-3
2.3 亲本成株期小种侵染量鉴定
通过半定量PCR方法对中抗、感病亲本及一些F1代杂交种的CYR23、CYR31、CYR33、CYR34侵染量进行鉴定,发现CYR33、CYR34在发病亲本中具有不同侵染量,CYR31在恢复系15CA50、不育系17L6078和15L7128上有少量侵染,没有检测到CYR23的侵染。随机选取表4中的F1代杂交种进行成株期菌量鉴定,发现亲本对不同小种的抗性在F1代杂交种上得到互补,例如15L7109中CYR33、CYR34侵染量较丰德存5号低,其F1代CYR34侵染量较丰德存5号明显降低。CYR34对不育系15L7109侵染量较西农807低,其F1代杂交种CYR34侵染量较西农807明显降低。恢复系川13品6、MR1101、川麦98及其F1代杂交种不发病,也没有检测到条锈菌侵染(图4)。
3 讨论
小麦抗病育种对小麦安全生产具有重要意义。MIEDANER等[37]将亲本和杂交种对小麦赤霉病的反应型进行相关性分析,发现亲本反应型的平均值与杂交种反应型有很大相关性,可以用来预测杂交种对赤霉病的抗性,这与本研究的结论一致;史丽丽等[16]在杂交小麦抗白粉病研究中利用成株期抗性的中国春与抗病材料杂交产生的F1代杂交种抗病性接近或略高于抗病亲本,其与感病品种杂交的F1代杂交种抗病性介于双亲之间,并且正反交实验结果为抗病性表现基本一致,表明杂交种抗性由双亲共同决定;Liu等[17]对1 749份小麦材料(15份感病父本,120份部分抗病母本,1 604份F1代杂交种,10份对照)进行多年多点的抗叶锈表型数据分析,发现源自抗病母本的F1代杂交种大多数抗病,总体反应型属于双峰分布,据此得出抗病的F1代杂交种有可能来自于特定亲本的假设。但根据本研究结果,认为更大的原因是部分双亲结合发生了抗性互补[38]。另外,许多材料在经过适当的组合和筛选后会在抗病方面有显著的超亲优势,双亲都显示高抗则F1代杂交种也高抗,双亲抗性有差异但抗性互补,F1代杂交种也能表现优良抗性。双亲抗性基因的显隐性与贡献都直接影响着F1代杂交种的抗性表现。通过对亲本及F1代杂交种成株期菌量的鉴定,发现F1代杂交种能够保持亲本的抗性,扩大对病原菌小种的抗谱范围。

1:ddH2O;2:Avocet S;3:Yr;4:17L9154;5:17L9160;6:17L9163;7:17L9210;8:17L9217;9:17L9195;10:17L6019;11:17L6062;12:17L6065;13:17L6067;14:17L6078;15:17L6085;16:17L7030;17:17L7106;18:17L7123;19:17L7140;20:17L9066;21:08L5070;22:15L7109;23:17L7128;24:17L7152

图3 亲本-F1代杂交种反应型的二元回归分析
CYR33对无毒性[39-40],本研究中含有的材料均未发现CYR33侵染。CYR34是2016年正式命名的新小种,其毒谱比CYR32、CYR33更广,并且在全国的小种流行趋势中所占比例逐年增加[39]。恢复系川14品16、川麦93、川麦98、MR1101、MY13-3、伟隆121和不育系17L9154、17L9163、17L9217、17L9066、08L5070等表现优良抗性,表明其中含有对CYR34未知的抗性基因,同时这些材料可以用于我国小麦抗条锈育种。CYR23对、、无毒性,对弱毒性[27],因此不育系和F1代杂交种中均未检测到该小种,但恢复系15CA50、丰德存5号、西农807未鉴定到抗性基因和CYR23侵染,表明这些材料含有抗CYR23的基因。当前小麦品种存在抗性与农艺性状协同性差的问题,农家品种蕴含优良的抗条锈病基因资源[41],但因综合农艺性状不佳,难以作为亲本直接用于小麦品种改良,因此在育种中好用并发挥重要作用的抗原极少,而具有优异农艺性状的材料往往抗病性不佳,只能达到中感或中抗水平。本研究发现部分不育系和恢复系可以互补,其F1代杂交种明显高于抗性中亲值或与最抗亲本相当,需要进一步对这些亲本的抗性基因进行鉴定,为今后的杂交小麦亲本选择提供帮助,快速、高效地培育新的杂交种应用于生产。

图4 CYR23、CYR31、CYR33、CYR34在植株中侵染量
培育持久抗病品种是小麦抗病育种的重要目标,培育早熟品种可以减少锈病对产量的影响,小麦的一些生理特征如气孔大小、蜡质层存在与否、角质层厚度、叶片角度和宽度等与决定持久抗性的特征非常相似[42]。育种过程中常用的选择方式有标记辅助选择、基因组选择以及表型选择等[43-44]。AGOSTINELLI等[45]通过表型选择和标记辅助选择的方法更大效率地提高了小麦杂交种对赤霉病的抗性;Salameh等[46]通过标记辅助选育叠加了两个抗赤霉病的QTL位点和,抗性比较结果为<≤+;Kim等[47]在杂交水稻抗病育种方面,利用标记辅助选育出多个抗白叶枯病、稻瘟病、褐飞虱并且农艺性状良好的杂交组合,为水稻的抗病虫育种提供了理论依据。与单个基因控制主要抗性相比,多基因型抗性基因的叠加可以达到持久抗性的目的,研究表明,一般叠加4—5个效应较大微效基因可达到接近免疫的水平,叠加2—3个达到中抗水平[48]。主效基因和微效基因有效结合也能产生良好抗性,如兼抗3种锈病及白粉病的多抗位点///、///和///与两个微效基因共同存在时会增加成株期抗性[49]。品种抗性基因单一和小种变异是导致小麦条锈病暴发的主要原因,例如由于来自于黑麦的1B/1R易位系抗病基因的大量使用曾导致某些地区小麦条锈病、白粉病大面积流行[50]。和分别在北方品系和川渝品系中有较高检出率,由于CYR34毒性较广,应减少对之前单一抗条锈基因的依赖。寻找新抗原迫在眉睫,分子标记能够精确地选择所需性状的材料,并且受环境的影响不大。没有与目的基因连锁的分子标记很难进行抗病基因累加,因此需加强抗病基因定位[51-52]。易感基因是使用基因组编辑工具进行抗性育种的重要目标,与抗性基因相比,有望为农作物提供更持久和更广谱的抗性[53-54]。
4 结论
F1代杂交种对条锈菌的反应型绝大多数趋于双亲反应型的平均值,因此可根据双亲反应型平均值预测F1代杂交种抗性。建议组合具有多个抗条锈病基因和对不同小种有抗性互补的亲本材料,以扩大F1代杂交种对条锈菌生理小种的抗性范围。应选择高产、优质、抗性良好的材料作为杂交种的亲本,才能获得适合生产上大面积推广应用的优势杂交组合。
[1] GODFRAY H C J, BEDDINGTON J R, CRUTE I R, HADDAD L, LAWRENCE D, MUIR J F, PRETTY J, ROBINSON S, THOMAS S M, TOULMIN C. Food security: the challenge of feeding 9 billion people., 2010, 327(5967): 812-818.
[2] FOLEY J A, RAMANKUTTY N, BRAUMAN K A, CASSIDY E S, GERBER J S, JOHNSTON M, MUELLER N D, O’CONNELL C, RAY D K, WEST P C, BALZER C, BENNETT E M, CARPENTER S R, HILL J, MONFREDA C, POLASKY S, ROCKSTROM J, SHEEHAN J, SIEBERT S, TILMAN D, ZAKS D P M. Solutions for a cultivated planet., 2011, 478(7369): 337-342.
[3] RAY D K, RAMANKUTTY N, MUELLER N D, WEST P C, FOLEY J A. Recent patterns of crop yield growth and stagnation.,2012, 3: 1293.
[4] RAY D K, MUELLER N D, WEST P C, FOLEY J A. Yield trends are insufficient to double global crop production by 2050., 2013, 8(6): e66428.
[5] WHITFORD R, FLEURY D, REIF J C, GARCIA M, OKADA T, KORZUN V, LANGRIDGE P. Hybrid breeding in wheat: technologies to improve hybrid wheat seed production., 2013, 64(18): 5411-5428.
[6] 丁位华, 冯素伟, 姜小苓, 王丹, 杨艳艳, 李婷婷, 茹振刚. 播期、密度和行距对BNS型杂交小麦光合及产量的影响. 麦类作物学报, 2017, 37(3): 366-375.
DING W H, FENG S W, JIANG X L, WANG D, YANG Y Y, LI T T, RU Z G. Effect of sowing date, density and row spacing on photosynthetic characteristic and yield of BNS hybrid wheat., 2017, 37(3): 366-375. (in Chinese)
[7] KOEMEL J E, GUENZI A C, CARVER B F, PAYTON M E, MORGAN G H, SMITH E L. Hybrid and pure line hard winter wheat yield and stability., 2004, 44(1): 107-113.
[8] KEMPE K, GILS M. Pollination control technologies for hybrid breeding., 2011, 27(4): 417-437.
[9] TUCKER E J, BAUMANN U, KOUIDRI A, SUCHECKI R, BAES M, GARCIA M, OKADA T, DONG C M, WU Y Z, SANDHU A, SINGH M, LANGRIDGE P, WOLTERS P, ALBERTSEN M C, CIGAN A M, WHITFORD R. Molecular identification of the wheat male fertility geneand its prospects for hybrid breeding., 2017, 8(1): 869.
[10] LONGIN C F, MÜHLEISEN J, MAURER H P, ZHANG H L, GOWDA M, REIF J C. Hybrid breeding in autogamous cereals., 2012, 125(6): 1087-1096.
[11] 朱冠楠, 曹幸穗. 杂交水稻和杂交小麦的选育(1960-2000年)—面向国民经济主战场的新中国农业科技. 中国科学院院刊, 2019, 34(9): 1036-1045.
ZHU G N, CAO X S. Breeding of hybrid rice and hybrid wheat (1960-2000)-China agricultural science and technology facing the main battlefield of national economy., 2019, 34(9): 1036-1045. (in Chinese)
[12] RASHEED A, XIA X C. From markers to genome-based breeding in wheat., 2019, 132(3): 767-784.
[13] CUI Y R, LI R D, LI G W, ZHANG F, ZHU T T, ZHANG Q F, ALI J, LI Z K, XU S D. Hybrid breeding of rice via genomic selection., 2020, 18(1): 57-67.
[14] 张杰, 董莎萌, 王伟, 赵建华, 陈学伟, 郭惠珊, 何光存, 何祖华, 康振生, 李毅, 彭友良, 王国梁, 周雪平, 王源超, 周俭民. 植物免疫研究与抗病虫绿色防控: 进展、机遇与挑战. 中国科学: 生命科学, 2019, 49(11): 1479-1507.
ZHANG J, DONG S M, WANG W, ZHAO J H, CHEN X W, GUO H S, HE G C, HE Z H, KANG Z S, LI Y, PENG Y L, WANG G L, ZHOU X P, WANG Y C, ZHOU J M. Plant immunity and sustainable control of pests in China: Advances, opportunities and challenges., 2019, 49(11): 1479-1507. (in Chinese)
[15] GUPTA P K, BALYAN H S, GAHLAUT V, SARIPALLI G, PAL B, BASNET B R, JOSHI A K. Hybrid wheat: past, present and future., 2019, 132(9): 2463-2483.
[16] 史丽丽, 张改生, 牛娜, 马守才, 李红霞, 徐开杰. 杂交小麦杂种一代白粉病抗性表现规律的研究. 麦类作物学报, 2009, 29(5): 919-924.
SHI L L, ZHANG G S, NIU N, MA S C, LI H X, XU K J. Performance on powdery mildew resistance of F1generation in hybrid wheat., 2009, 29(5): 919-924. (in Chinese)
[17] LIU F, ZHAO Y S, Beier S, Jiang Y, Thorwarth P, HLongin C F, Ganal M, Himmelbach A, Reif J C, Schulthess A w. Exome association analysis sheds light onto leaf rust () resistance genes currently used in wheat breeding (L.)., 2019, doi: 10.1111/pbi.13303.
[18] BOEVEN P H, WURSCHU T, WEISSMANN S, MIEDANER T, MAURER H P. Prediction of hybrid performance for Fusarium head blight resistance in triticale (×Wittmack)., 2016, 207(3): 475-490.
[19] ALI N, HESLOP-HARRISON J P, AHMAD H, GRAYBOSCH R A, HEIN G L, SCHWARZACHER T. Introgression of chromosome segments from multiple alien species in wheat breeding lines with wheat streak mosaic virus resistance., 2016, 177(2): 114-123.
[20] 赵仁慧, 刘炳亮, 寿路路, 陈甜甜, 王海燕, 王秀娥, 别同德. 分子标记辅助聚合抗小麦黄花叶病和白粉病育种. 麦类作物学报, 2017, 37(12): 1541-1549.
ZHAO R H, LIU B L, SHOU L L, CHEN T T, WANG H Y, WANG X E, BIE T D. Pyramiding disease resistance to wheat yellow mosaic virus and powdery mildew by molecular marker-assisted selection., 2017, 37(12): 1541-1549. (in Chinese)
[21] 崔彩红, 房伟强, 李兴锋, 王洪刚, 鲍印广. 携带抗秆锈病基因、小麦新种质的分子标记辅助选育. 山东农业科学, 2015, 47(4): 13-17.
CUI C H, FANG W Q, LI X F, WANG H G, BAO Y G. Marker assisted selection of wheat germplasms with stem rust resistance genesand., 2015, 47(4): 13-17. (in Chinese)
[22] CHEN W Q, Wellings C, CHEN X M, KANG Z S, LIU T G. Wheat stripe (yellow) rust caused byf. sp.., 2014, 15(5): 433-446.
[23] 康振生, 王晓杰, 赵杰, 汤春蕾, 黄丽丽. 小麦条锈菌致病性及其变异研究进展. 中国农业科学, 2015, 48(17): 3439-3453.
KANG Z S, WANG X J, ZHAO J, TANG C L, HUANG L L. Advances in research of pathogenicity and virulence variation of the wheat stripe rust fungusf. sp., 2015, 48(17): 3439-3453. (in Chinese)
[24] 张玉薇, 刘太国, 刘博, 高利, 陈万权. 中国75个国审小麦品种抗条锈基因推导. 植物保护学报, 2014, 41(1): 45-53.
ZHANG Y W, LIU T G, LIU B, GAO L, CHEN W Q. Gene postulation of stripe rust resistance genes of 75 Chinese commercial wheat cultivars., 2014, 41(1): 45-53. (in Chinese)
[25] 李中安. 一种以蓝粒为标记性状的两系法杂交小麦的选育方法: CN200610042629.8[P]. (2006-09-06)[2019-12-17].
LI Z A. A breeding method of two-line hybrid wheat marked by blue grain: CN200610042629.8[P]. (2006-09-06)[2019-12-17]. (in Chinese)
[26] MARCHAL C, ZHANG J P, ZHANG P, FENWICK P, STEUERNAGEL B, ADAMSKI N M, BOYD L, MCLNTOSH R, WULFF B B H, BERRY S, LAGUDAH E, UAUY C. BED-domain-containing immune receptors confer diverse resistance spectra to yellow rust., 2018, 4(9): 662-668.
[27] ZENG Q D, HAN D J, WANG Q L, YUAN F P, WU J H, ZHANG L, WANG X J, HAUNG L L, CHEN X M, KANG Z S. Stripe rust resistance and genes in Chinese wheat cultivars and breeding lines., 2014, 196(2): 271-284.
[28] KLYMIUK V, YANIV E, HUANG L, RAATS D, FATIUKHA A, CHEN S,Feng L, Frenkel Z, Krugman T, Lidzbarsky G, Chang W, Jääskeläinen M J, Schudoma C, Paulin L, Laine P, Bariana H, Sela H, Saleem K, Sørensen C K, Hovmøller M S, Distelfeld A, Chalhoub B, Dubcovsky J, Korol A B, Schulman A H, Fahima T. Cloning of the wheatresistance gene sheds light on the plant tandem kinase-pseudokinase family., 2018, 9(1): 3735.
[29] YANG Y H, CHEN F J, HAN D J, RUAN R W, LI B Q, YU Y, BI C W. Evaluation of resistance of current wheat cultivars and breeding lines to stripe rust from three Gorges reservoir area., 2017, 83(5): 283-290.
[30] 李北, 徐琪, 杨宇衡, 王琪琳, 曾庆东, 吴建辉, 穆京妹, 黄丽丽, 康振生, 韩德俊. 重庆麦区小麦品种(系)抗条锈性评价与基因分析. 中国农业科学, 2017, 50(3): 413-425.
LI B, XU Q, YANG Y H, WANG Q L, ZENG Q D, WU J H, MU J M, HUANG L L, KANG Z S, HAN D J. Stripe rust resistance and genes in chongqing wheat cultivars and lines.,2017, 50(3): 413-425. (in Chinese)
[31] CHOQUER M, BOCCARA M, VIDAL-CROS A. A semi- quantitative RT-PCR method to readily compare expression levels withinmultigenic familiesand., 2003, 43(4): 303-309.
[32] XU Q, TANG C L, WANG L K, ZHAO C C, KANG Z S, WANG X J. Haustoria-arsenals during the interaction between wheat andf. sp.., 2019, doi: 10.1111/mpp.12882.
[33] 曹丽华, 康振生, 赵杰, 黄丽丽, 魏国荣. 中国小麦条锈菌4个流行小种的RAPD标记. 西北农林科技大学学报(自然科学版), 2004, 32(7): 37-40.
CAO L H, KANG Z S, ZHAO J, HUANG L L, WEI G R. RAPD markers off. sp.in China., 2004, 32(7): 37-40. (in Chinese)
[34] 曹丽华, 康振生, 郑文明, 黄丽丽, 李振岐. 小麦条锈菌条中31号生理小种SCAR检测标记的建立. 菌物学报, 2005, 24(1): 98-103.
CAO L H, KANG Z S, ZHENG W M, HUANG L L, LI Z Q. The development of SCAR detection marker off. sp.race CYR31 in China., 2005, 24(1): 98-103. (in Chinese)
[35] WANG B T, HU X P, LI Q, HAO B J, ZHANG B, LI G B, KANG Z S. Development of race-specific SCAR markers for detection of Chinese races CYR32 and CYR33 off. sp.., 2010, 94(2): 221-228.
[36] 郭婧, 李敏州, 夏滔, 李高宝, 申雪雪, 樊玉, 李强, 王保通. 小麦条锈菌新菌系V26的SCAR 检测标记. 植物病理学报, 2014, 44(5): 449-454.
GUO J, LI M Z, XIA T, LI G B, SHEN X X, FAN Y, LI Q, WANG B T. Race-specific SCAR marker for new strain V26 off. sp.in China., 2014, 44(5): 449-454. (in Chinese)
[37] MIEDANER T, SCHULTHESS A W, GOWDA M, REIF J C, LONGIN C F H. High accuracy of predicting hybrid performance of Fusarium head blight resistance by mid-parent values in wheat., 2017, 130(2): 461-470.
[38] 吴春太, 徐如宏, 张庆勤. 高产抗病优质互补的品种间杂交研究. 西南农业学报, 2010, 23(6): 1891-1894.
WU C T, XU R H, ZHANG Q Q. Study of hybridization between varieties with exchangeable supply in high-yield disease resistance and good quality., 2010, 23(6): 1891-1894. (in Chinese)
[39] 刘博, 刘太国, 章振羽, 贾秋珍, 王保通, 高利, 彭云良, 金社林, 陈万权. 中国小麦条锈菌条中34号的发现及其致病特性. 植物病理学报, 2017, 47(5): 681-687.
LIU B, LIU T G, ZHANG Z Y, JIA Q Z, WANG B T, GAO L, PENG Y L, JIN S L, CHEN W Q. Discovery and pathogenicity of CYR34, a new race off. sp.in China.,2017, 47(5): 681-687. (in Chinese)
[40] 姚强, 王洁荣, 孟岩, 詹刚明, 黄丽丽, 康振生. 中国小麦条锈病菌CYR32和CYR33的毒性及基因型多样性. 植物保护学报, 2018, 45(1): 46-52.
YAO Q, WANG J R, MENG Y, ZHAN G M, HUANG L L, KANG Z S. Virulence and genotypic diversity of wheat stripe rust races CYR32 and CYR33 in China., 2018, 45(1): 46-52. (in Chinese)
[41] 黄苗苗, 孙振宇, 曹世勤, 贾秋珍, 刘太国, 陈万权. 223份小麦农家品种田间抗条锈病性评价及抗病基因分子检测. 植物保护学报, 2018, 45(1): 90-100.
HUANG M M, SUN Z Y, CAO S Q, JIA Q Z, LIU T G, CHEN W Q. Evaluation of the resistance of 223 wheat landraces in Gansu Province to stripe rust and molecular detection., 2018, 45(1): 90-100. (in Chinese)
[42] REHMAN A U, SAJJAD M, KHAN S H, AHMAD N. Prospects of wheat breeding for durable resistance against brown, yellow and black rust fungi., 2013, 15(6): 1209-1220.
[43] Liu G Z, Zhao Y S, GOWDA M, LONGIN C F, REIF J C, METTE M F. Predicting hybrid performance for quality traits through genomic-assisted approaches in Central European wheat., 2016, 11(7): e0158635.
[44] STEINER B, BUERSTMAYR M, MICHEL S, SCHWEIGER W, LEMMENS M, BUERSTMAYR H. Breeding strategies and advances in line selection for Fusarium head blight resistance in wheat., 2017, 42(3): 165-174.
[45] AGOSTINELLI A M, CLARK A J, BROWN-GUEDIRA G, VAN Sanford D A. Optimizing phenotypic and genotypic selection for Fusarium head blight resistance in wheat., 2012, 186(1): 115-126.
[46] SALAMEH A, BUERSTMAYR M, STEINER B, NEUMAYER A, LEMMENS M, BUERSTMAYR H. Effects of introgression of two QTL for fusarium head blight resistance from Asian spring wheat by marker-assisted backcrossing into European winter wheat on fusarium head blight resistance, yield and quality traits., 2010, 28(4): 485-494.
[47] KIM M S, OUK S, JUNG K H, SONG Y, YANG J Y, CHO Y G. Breeding hybrid rice with genes resistant to diseases and insects using marker-assisted selection and evaluation of biological assay., 2019, 7(3): 272-286.
[48] SINGH R P, HUERTA-ESPINO J, WILLIAM H M. Genetics and breeding for durable resistance to leaf and stripe rusts in wheat., 2005, 29(2): 121-127.
[49] Singh R P, Singh P K, Rutkoski J, Hodson D P, He X, Jorgensen L N, Hovmoller M S, Huerta-Espino J. Disease impact on wheat yield potential and prospects of genetic control., 2016, 54: 303-322.
[50] 李峰奇, 韩德俊, 魏国荣, 曾庆东, 黄丽丽, 康振生. 黄淮麦区126 个小麦品种(系)抗条锈病基因的分子检测. 中国农业科学, 2008, 41(10): 3060-3069.
LI F Q, HAN D J, WEI G R, ZENG Q D, HUANG L L, KANG Z S. Molecular detection of stripe rust resistant genes in 126 winter wheat varieties from the Huanghuai wheat region., 2008, 41(10): 3060-3069. (in Chinese)
[51] 任勇, 李生荣, 周强, 杜小英, 何员江, 魏育明, 郑有良. 134份四川小麦品种(系)的条锈病抗性评价. 麦类作物学报, 2014, 34(6): 847-853.
REN Y, LI S R, ZHOU Q, DU X Y, HE Y J, WEI Y M, ZHENG Y L. Evaluation of resistance to stripe rust of 134 wheat cultivars and lines from Sichuan province., 2014, 34(6): 847-853. (in Chinese)
[52] SAVADI S, PRASAD P, KASHYAP P L, BHARDWAJ S C. Molecular breeding technologies and strategies for rust resistance in wheat () for sustained food security., 2018, 67(4): 771-791.
[53] WANG Y P, CHENG X, SHAN Q W, ZHANG Y, LIU J X, CAO C X, QIU J L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew., 2014, 32(9): 947-951.
[54] ARAKI M, ISHII T. Towards social acceptance of plant breeding by genome editing., 2015, 20(3): 145-149.
The Resistance Prediction of Wheat Hybrids based on the Sensibility of their Parents to Stripe Rust
ZHOU TianYu1, LI JiangLing1, YANG Lan1, RUAN RenWu2, YANG YuHeng3, LI ZhongAn1
(1Citrus Research Institute, Southwest University, Chongqing 400712;2College of Agronomy and Biotechnology, Southwest University, Chongqing 400716;3College of Plant Protection, Southwest University, Chongqing 400716)
【】The objective of this study is to predict the resistance of wheat hybrids based on the resistance evaluation of their parental lines to stripe rust, and to enhance the predictability of disease resistance breeding in hybrid wheat.【】A sensitive wheat variety, Mingxian 169, was used as negative control. A total of 13 restorer lines (male parents), 21 sterile lines (female parents), and their F1hybrids were inoculated with a suspension of a mixture of fresh urediniospores (equal amounts of stripe rust pathogen physiological races CYR23, CYR31, CYR33, and CYR34)in the field. Stripe rust resistance genes,,,,,andwere identified by PCR using their gene or linked molecular markers.In addition, semi-quantitative PCR was used to quantify the different stripe rust races in the adult plants of parent lines and a part of F1hybrids.【】,andwere not identified in all materials,was mostly existed in Sichuan wheat lines,andwere mostly existed in the northern lines,was not identified in restorer lines. The parental resistance genes were polymerized in F1hybrids, which is consistent with the genetic law, indicating that the molecular marker can be used in wheat assistant breeding. The restorer lines from Sichuan and their F1hybrids showed high resistance against stripe rust, speculating that there are homozygous dominant resistance genes against stripe rust. Meanwhile, these wheat lines can be used for stripe rust resistance breeding in China. The infection types (ITs) of F1hybrids tended to the average of parental infection types. The results of binary regression analysis showed a significant correlation between parent lines and F1hybrids in ITs (2=0.812). Althoughandresistance genes were absent in all tested wheat lines, the restorers from Sichuan Province and their F1hybrids showed higher resistance, speculating that those lines might carry unknown resistance genes against CYR34. The semi-quantitative PCR results indicated that CYR23 was not detected in all wheat lines. Only the restorer line 15CA50, sterile lines 17L6078 and 15L7128 were infected with a small amount of CYR31. The tested restorer lines Chuan 13 pin 6, MR1101, Chuanmai 98 and their F1hybrids were not infected by CYR33 or CYR34. Meanwhile, the parents with complementary resistance to different races were found to increase the resistance level of F1hybrids effectively.【】The F1hybrids resistance to strip rust can be predicted according to the average of the ITs of parental lines, the higher levels of disease resistance of parents, the better resistance of F1hybrids. Wheat varieties or lines with complementary resistance to different races of stripe rust should be screened as parents to improve the resistance level of F1hybrids. The results revealed the rule of disease resistance between parents and their F1hybrids, as well as providing a practical strategy for hybrid wheat disease resistance breeding.
hybrid wheat; stripe rust; resistance prediction;f. sp.; physiological race

10.3864/j.issn.0578-1752.2020.09.009
2019-12-17;
2020-02-12
国家重点研发计划(2016YFD0101603,2018YFD0200500)、国家自然科学基金(31801719)
周天宇,E-mail:1522619059@qq.com。通信作者李中安,E-mail:zhongan@cric.cn。通信作者杨宇衡,E-mail:yyh023@swu.edu.cn
(责任编辑 岳梅)
