羊口疮病毒B2L、F1L 基因融合真核表达质粒诱导小鼠的免疫应答及IL-2 对其作用影响的研究
2020-06-01鲜思美饶体宇吴伯梅包涛涛闵德省包细明
鲜思美,张 友,李 婷,杨 倩,饶体宇,吴伯梅,包涛涛,闵德省,包细明,张 益
(1. 贵州大学 动物科学学院,贵州 贵阳 550025;2. 贵州省动物疫病研究所,贵州 贵阳 550025)
羊传染性脓疱病由羊口疮病毒(Orf virus,OrfV)所致的一种人兽共患传染病[1-2],该病传染性强,发病率高,以羔羊最为易感;病羊口唇等部位的皮肤及粘膜出现红斑、丘疹、脓疮、溃疡是Orf的常见症状[3-4]。用于预防Orf 的疫苗主要有灭活苗和弱毒疫苗,均能在一定程度上降低疾病的感染率,但是灭活苗不能有效激发细胞免疫,弱毒疫苗存在毒力返强等潜在的风险。随着规模化养羊业的快速发展,Orf 的危害越发严重,因此,利用基因工程技术研发一种新型疫苗对防治Orf 具有重大意义[5]。DNA 疫苗作为一种新型疫苗,没有毒力返强,同时具有制备简单、成本低廉、质粒性质稳定、便于运输和保存等优点。
OrfV 的B2L 和F1L 基因是两个重要的免疫原性基因,可以诱导机体产生良好的免疫反应。冯平等通过将B2L 和F1L 基因串联,并在昆虫杆状病毒表达系统获得了部分分泌型的B2L 和F1L 串联重组蛋白[6]。赵魁构建了pCDNA3.1-ORFV 011(B2L)、pCDNA3.1-ORFV 059(F1L)和pCDNA3.1-ORFV 011(B2L)-linker-ORFV(F1L)质粒,通过免疫小鼠表明了pCDNA3.1-ORFV 011(B2L)-linker-ORFV(F1L)质粒免疫效果优于pCDNA3.1-ORFV 011(B2L)、pCDNA3.1-ORFV 059(F1L)质粒[7]。本实验室前期构建了真核表达 质 粒pVAX1-IL-2、pVAX1-F1L 和pVAX1-B2L,并通过免疫小鼠证明其具有良好的免疫原性[8]。在此基础上,本研究构建了OrfV pVAX1-B2L-F1L 融合基因真核表达质粒,并以pVAX1-IL-2 为细胞因子佐剂免疫小鼠,评价其诱导小鼠的免疫应答效果。以期为后续试验核酸疫苗PVAX1-B2L-F1L 免疫宿主动物奠定基础。
1 材料与方法
1.1 质粒及实验动物 真核表达质粒pVAX1-IL-2、pVAX1-B2L-F1L 由贵州省动物疫病研究室构建、鉴定并保存;OrfV 阳性高免兔血清、MDBK 细胞由本实验室保存;165 只5 周龄~6 周龄KM 系小鼠,雌雄各半,体质量18 g~20 g。购自中国人民解放军第三军医大学实验动物中心。
1.2 主要试剂 RNA 提取试剂盒、HiFiScript cDNA第一链合成试剂盒购自北京康为世纪生物科技有限公司;脂质体2000(Lipofatamiane 2000)购自美国Invitrogen 公司;FITC 标记的羊抗兔IgG 购自北京博奥森生物技术有限公司;DMSO、ConA、MTT 购自美国Solarbio 公司;无内毒素质粒小提中量试剂盒、无内毒素质粒大提试剂盒、红细胞裂解液购自天根深化科技(北京)有限公司;OrfV 抗体(OrfV-Ab)酶联免疫试剂盒购自上海酶联生物科技有限公司;小鼠IL-2、IFN-γ、IL-4 和IL-6 ELISA 定量检测试剂盒购自武汉基因美公司。
1.3 融合基因真核表达质粒pVAX1-B2L-F1L 在MDBK 细胞中的表达 参照Lipofectamine 2000 说明书,将pVAX1-B2L-F1L、pVAX1 质粒分别转染MDBK 细胞,48 h 后收集转染的细胞;采用RT-PCR 从mRNA 水平检测pVAX1-B2L-F1L 转录情况[9];以OrfV 阳 性 高 免 兔 血 清(1∶50)1 为 一 抗;羊 抗 兔IgG-FITC(1∶500)为二抗,采用间接免疫荧光试验(IFA)检 测pVAX1-B2L-F1L 在MDBK 细 胞 中 的 表达。同时以pVAX1 空载体和正常细胞作为对照。
1.4 小鼠实验设计 将160只小鼠随机分成4组,40只/组,采用腿部肌肉注射途径免疫重组质粒,各组免疫的质粒与剂量见表1。间隔一周以相同剂量、途径加强免疫,共免疫3 次。免疫后4 组小鼠每周眼球采血,每组每周采5 只小鼠,共采血8 周,分离血清,-20 ℃保存。眼球采血后,颈椎脱臼法迫杀小鼠,无菌采集其脾脏,用于小鼠脾细胞增殖试验。
1.5 免疫小鼠血清中特异性抗体水平检测 参照小鼠OrfV-Ab ELISA 试剂盒说明书检测各组小鼠首免后不同时间段小鼠血清中OrfV 特异性抗体(该试剂盒中,当检测样品浓度<2 pg/mL 时,判为阴性)。所得数据用SPSS 软件进行统计学分析。
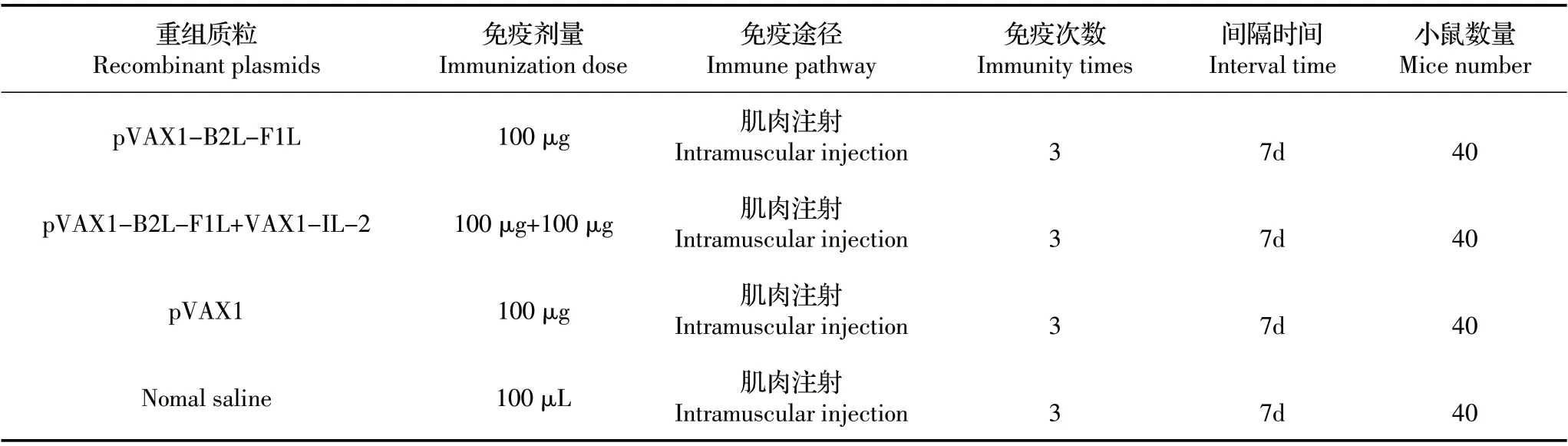
表1 小鼠实验设计Table 1 Mouse experimental design
1.6 免疫小鼠脾淋巴细胞增殖试验 每周每组取5只小鼠眼球采血后断颈迫杀小鼠,酒精浸泡5 min,制备脾淋巴细胞悬液:将无菌采集的脾脏放入有1640 的无菌EP 管中,将其剪碎,用200 目细胞筛研磨制成细胞悬液,室温离心10 min,弃上清;加入2 mL 无菌红细胞裂解液重悬细胞沉淀,37 ℃放置15 min 后,室温离心10 min,弃上清;加入1640 重悬细胞沉淀,室温离心10 min;加入含10%胎牛血清的1640 培养液调整细胞数达1×106个/mL。用终浓度为50 μg/mL 的ConA 诱导,采用MTT 法[9]检测免疫小鼠脾淋巴细胞增殖情况。酶标仪测定各孔OD490nm值。所得数据用SPSS 软件进行统计学分析。
1.7 免疫小鼠血清中Th1/ Th2 型细胞因子检测将各组小鼠首免后不同时间段分离的血清,通过ELISA 方法测定小鼠血清中Th1 型(IL-2、IFN-γ)、Th2 型(IL-4、IL-6)细胞因子含量。所得数据用SPSS 软件进行统计学分析。
2 结 果
2.1 融合基因转录水平和表达的检测结果 提取细胞总RNA,通过RT-PCR 方法检测B2L-F1L 融合基因的转录水平。结果显示,转染pVAX1-B2L-F1L的细胞经PCR 扩增出约2 181 bp 的目的条带,与预期一致。但pVAX1 转染的细胞和未转染的细胞检测结果为阴性(图1);通过间接免疫荧光(IFA)检测pVAX1-B2L-F1L 重组质粒的表达产物。结果显示,转染重组质粒pVAX1-B2L-F1L 的细胞可见绿色荧光,同形对照组[8](不加一抗,只加二抗)转染的细胞未见绿色荧光,转染pVAX1 空载体的细胞和正常细胞也未见绿色荧光(图2)。表明融合基因B2L-F1 能够在MDBK 细胞中表达。

图1 B2L-F1L 基因mRNA 在MDBK 细胞中的转录水平Fig.1 mRNA transcription of eukaryotic expression plasmid pVAX1-B2L-F1L in MDBK cells
2.2 免疫小鼠血清特异性抗体水平检测结果 采用ELISA方法检测小鼠血清中OrfV特异性抗体。结果显示,pVAX1-B2L-F1L 免疫组和pVAX1-B2L-F1L+pVAX1-IL-2 免疫组小鼠在首免后第1 周迅速产生抗体,两组特异性抗体水平均在首免后第5 周时达到最高;首免后第1~8周,pVAX1-B2L-F1L+pVAX1-IL-2 免疫组小鼠产生OrfV 特异性抗体水平均高于pVAX1-B2L-F1L免疫组,其中首免后第4~6周pVAX1-B2L-F1L+pVAX1-IL-2免疫组小鼠OrfV特异性抗体水平极显著高于pVAX1-B2L-F1L 免疫组(p<0.01),而pVAX1 组和生理盐水对照组小鼠自始至终都没有产生OrfV 特异性抗体(图3)。结果表明,融合基因真核表达质粒pVAX1-B2L-F1L 能够有效刺激小鼠机体产生OrfV 特异性抗体,且pVAX1-IL-2 作为免疫佐剂能够有效增强pVAX1-B2L-F1L的体液免疫应答水平。
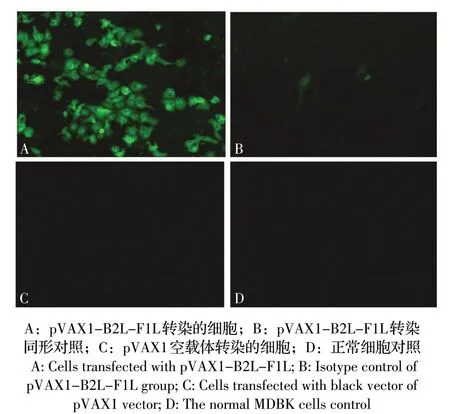
图2 融合基因表达的IFA 检测(400×)Fig.2 IFA test results(400×)

图3 免疫小鼠血清中OrfV 特异性抗体动态变化Fig.3 Dynamic variation of specific antibodies against OrfV in serum of immunized mice
2.3 免疫小鼠脾淋巴细胞增殖试验结果 对免疫后不同时间段迫杀的小鼠取脾经分离培养小鼠脾淋巴细胞进行非特异性刺激后,采用MTT 法检测其增殖情况。结果显示:首免后第2 周~第8 周,pVAX1-B2L-F1L 免疫组与pVAX1-B2L-F1L+pVAX1-IL-2 免疫组小鼠脾淋巴细胞OD490nm值均极显著高于pVAX1 组和生理盐水(p<0.01)组;pVAX1-B2L-F1L+pVAX1-IL-2 免疫组小鼠脾淋巴细胞OD490nm值高于pVAX1-B2L-F1L免疫组,其中首免后第2周和第6周,前者脾淋巴细胞OD490nm值极显著高于后者(p<0.01),首免后第5 周二者差异显著(p<0.05)(图4)。结果表明,pVAX1-B2L-F1L 真核表达质粒免疫小鼠后能够刺激小鼠脾淋巴细胞增殖,且pVAX1-IL-2 能够促进pVAX1-B2L-F1L 质粒诱导的小鼠脾淋巴细胞的增殖。
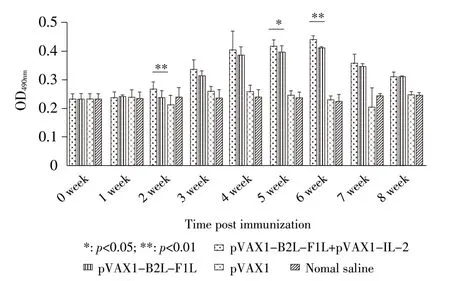
图4 免疫小鼠脾淋巴细胞增殖动态变化Fig.4 Dynamic variation of splenic lymphocyte proliferation in immunized mice
2.4 免疫小鼠血清中Th1 型细胞因子的检测结果
2.4.1 免疫小鼠血清中IL-2 的检测结果 采用ELISA 方法检测各组小鼠不同时间段血清中IL-2 的含量。结果显示,pVAX1-B2L-F1L 免疫组和pVAX1-B2L-F1L+pVAX1-IL-2 免疫组小鼠在首免后第1 周即开始迅速分泌IL-2,在首免后第1 周~第8 周前者小鼠IL-2的分泌量均比后者的分泌量高,其中首免后第1周二者之间差异显著(p<0.05);首免后第2周~第7周二者差异均极显著(p<0.01);在首免后第8 周时二者差异不显著(p>0.05);与pVAX1 空载体和生理盐水对照组相比,在首免后第1 周~第8 周二者差异极显著(p<0.01)(图5)。结果表明,pVAX1-IL-2 能够促进pVAX1-B2L-F1L 诱导的小鼠机体中IL-2 的分泌。
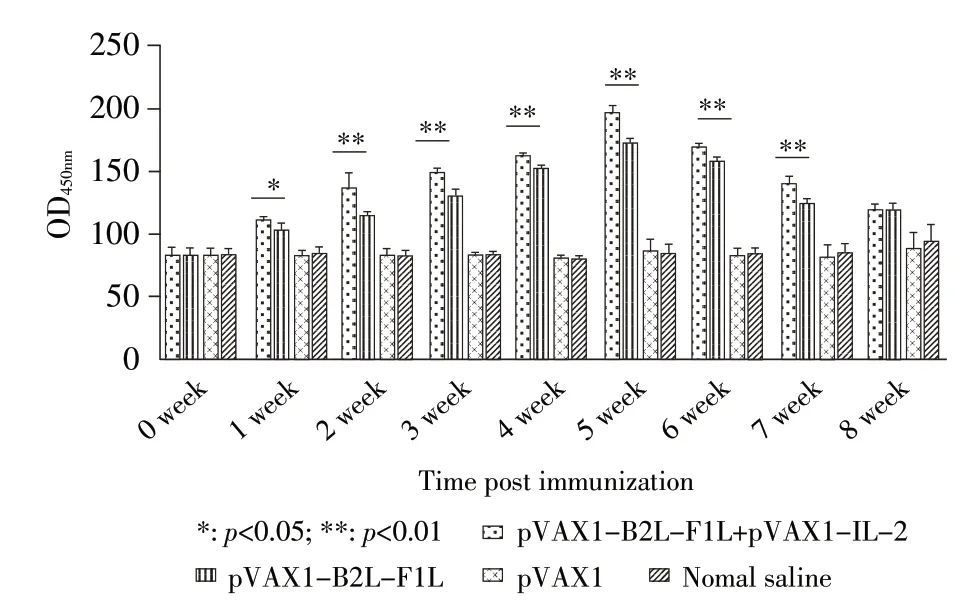
图5 免疫小鼠血清中IL-2 动态变化Fig.5 Dynamic variation of serum IL-2 in immunized mice
2.4.2 免疫小鼠血清中IFN-γ的检测结果 采用ELISA 方法检测各组小鼠不同时间段血清中IFN-γ 的含量。结果显示,在首免后第1 周~第8 周,pVAX1-B2L-F1L 免 疫 组 和pVAX1-B2L-F1L+pVAX1-IL-2 免疫组小鼠血清中分泌IFN-γ 的量均极显著高于pVAX1 组和生理盐水组(p<0.01);首免后第1 周pVAX1-B2L-F1L 免疫组小鼠IFN-γ分泌量显著高于pVAX1-B2L-F1L+pVAX1-IL-2 免疫组(p<0.05),首免后第2 周二者之间差异不显著,但首免后第3 周~第8 周后者IFN-γ分泌量极显著高于前者(p<0.01)(图6)。结果表明,pVAX1-IL-2 能够促进pVAX1-B2L-F1L 诱导小鼠机体中IFN-γ的分泌。
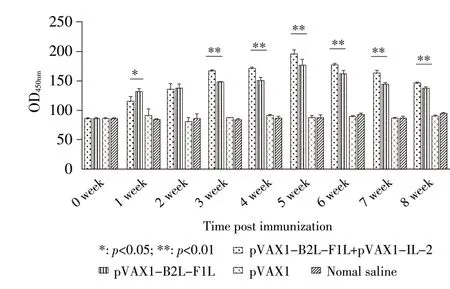
图6 免疫小鼠血清中IFN-γ动态变化Fig.6 Dynamic variation of serumIFN-γ in immunized mice
2.5 免疫小鼠血清中Th2 型细胞因子的检测结果
2.5.1 免疫小鼠血清中IL-4 的检测结果 采用ELISA 方法检测各组小鼠不同时间段血清中IL-4 的含量。结果显示,pVAX1-B2L-F1L 免疫组和pVAX1-B2L-F1L+pVAX1-IL-2 免疫组首免后1 周内能够刺激小鼠机体迅速分泌IL-4,首免后第1 周~第6 周二者均极显著高于pVAX1 组和生理盐水组(p<0.01),首免后第7 周4 个组小鼠IL-4 的分泌量差异不显著(p>0.05);首 免 后 第1 周、2 周、3 周、6 周pVAX1-B2L-F1L+pVAX1-IL-2 免疫组小鼠IL-4 分泌量均极显著或显著高于pVAX1-B2L-F1L 免疫组(p<0.01;p<0.05);在首免后第4 周、5 周、8 周后者小鼠血清中IL-4 分泌量高于前者,其中首免后第4 周和第8 周 二 者 差 异 显 著(p<0.05)(图7)。结 果 表 明,pVAX1-B2L-F1L+pVAX1-IL-2 免疫和pVAX1-B2LF1L 免疫均够能刺激小鼠机体产生IL-4,但pVAX1-IL-2 对pVAX1-B2L-F1L 诱导小鼠机体分泌IL-4 的效果不显著。
2.5.2 免疫小鼠血清中IL-6 检测结果 采用ELISA方法检测各组小鼠不同时间段血清中IL-6 的含量。结果显示,pVAX1-B2L-F1L 免疫组和pVAX1-B2LF1L+pVAX1-IL-2 免疫组小鼠首免后1 周内均能够刺激小鼠机体快速分泌IL-6,首免后第1 周、2 周、3周、6 周、7 周二者分泌IL-6 水平均极显著高于pVAX1 空载体组和生理盐水对照组(p<0.01);在首免后第1 周、2 周,pVAX1-B2L-F1L 免疫组小鼠血清中IL-6 的分泌量极显著高于pVAX1-B2L-F1L+pVAX1-IL-2 免 疫 组(p<0.01),首 免 后 第4 周、5周、8 周前者分泌IL-6 量也高于后者,但是二者之间差异不显著(p>0.05);在首免后第3 周、6 周、7周,pVAX1-B2L-F1L+pVAX1-IL-2 免疫组小鼠血清中IL-6 的分泌量高于pVAX1-B2L-F1L 免疫组,但二者之间差异不显著(p>0.05)(图8)。结果表明,pVAX1-B2L-F1L+pVAX1-IL-2 免疫和pVAX1-B2LF1L 免疫均能够刺激小鼠机体产生IL-6,但pVAX1-IL-2 对pVAX1-B2L-F1L 诱导小鼠机体分泌的IL-6无明显作用。
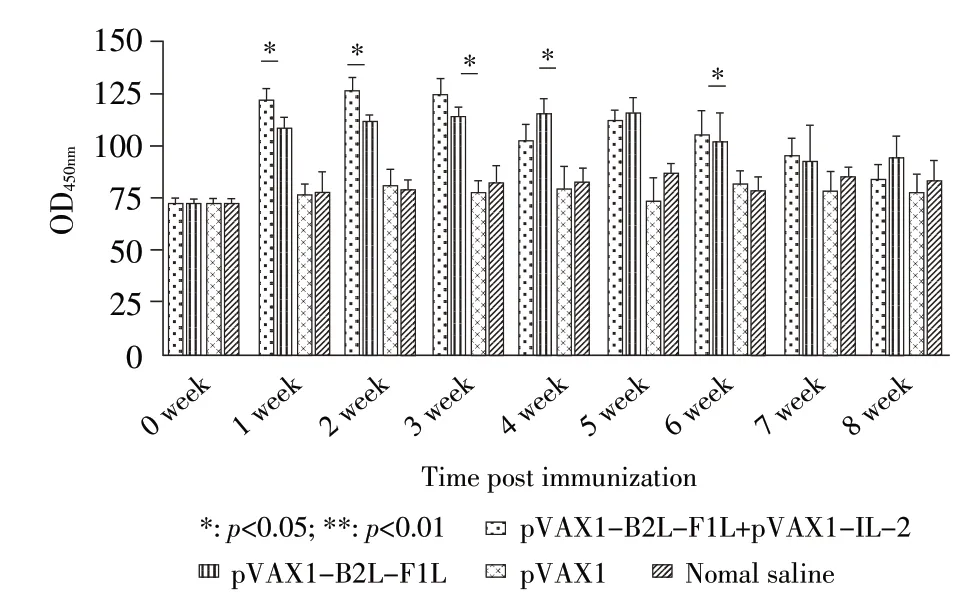
图7 免疫小鼠血清中IL-4 动态变化Fig.7 Dynamic variation of IL-4 in sera of immunized mice

图8 免疫小鼠血清中IL-6 动态变化Fig.8 Dynamic variation of IL-6 in sera of immunized mice
3 讨 论
Orf 呈世界性分布,给养羊业造成重大经济损失,严重阻碍畜牧业发展。OrfV不仅能感染羊,近年来也有关于其感染猫和家养驯鹿的报道[10];同时也对人类健康造成威胁,具有重要的公共卫生学意义[11]。国内外对Orf 的研究已经证明OrfV 是一种变异性较强的病毒,各地分离的病毒在致病性上均存在着较大差异[12]。OrfV 感染动物后,机体能产生很强的免疫反应,包括中性粒细胞、皮肤树突细胞、T 细胞和B 细胞、CD4+T 细胞和少量的CD8+T 细胞产生的抗病毒作用。OrfV 具有高度的嗜上皮性,介导的是细胞免疫应答,体液免疫应答水平低下,并且主要以局部免疫为主[13-14]。
DNA 疫苗具有能表达天然蛋白抗原,主要诱导机体产生细胞免疫应答的特点。外源性抗原进入机体后,与B 细胞膜上的SIg 特异性结合,活化B 细胞,启动体液免疫应答。由于激活B 细胞,需要经过细胞分化、增殖等过程,所以初次免疫应答产生抗体水平较低;再次加强免疫后,B 细胞分泌的抗体量增加,且分泌快速[15]。本研究通过ELISA 检测免疫前后不同时间段小鼠血清中OrfV 特异性抗体水平,结果表明,pVAX1-B2L-F1L 能够有效刺激小鼠机体产生OrfV 特异性抗体,且加强免疫后小鼠血清中OrfV 特异性抗体水平开始升高,在首免后第5 周达到峰值,第6 周开始抗体水平缓慢下降,但是在第8 周时的抗体水平仍然为阳性,这一检测结果与赵魁构建的质粒pCDNA3.1-OrfV011(B2L)-linker-OrfV059(F1L)免疫小鼠产生抗体水平动态一致[16]。利用ConA 对不同时间段分离培养的小鼠脾淋巴细胞进行非特异性刺激,MTT 法检测其增殖情况,结果与前期研究的pVAX1-B2L、pVAX1-F1L 单独免疫小鼠后的脾淋巴增殖试验相比,在首免后第1 周~第8 周,pVAX1-B2L-F1L 免疫组小鼠脾淋巴细胞OD490nm值 显 著 高 于pVAX1-B2L[17]、pVAX1-F1L 免 疫组[18],表明OrfV 双基因融合真核表达质粒pVAX1-B2L-F1L 促进小鼠脾淋巴细胞增殖的能力高于单基因真核表达质粒。
效应T 细胞主要有辅助性T 细胞(Th)和细胞毒性T 细胞(Tc)两大类,Th 细胞根据功能分为两个亚群Th1 和Th2,Th1 细 胞 主 要 分 泌IL-2、IFN-γ、IL-12 和IL-18 等细胞因子,主要介导细胞免疫应答;Th2 细胞主要分泌IL-4、IL-6 和IL-10 等细胞因子,主要促进B 细胞增殖、分化成浆细胞,分泌特异性抗体,介导体液免疫应答[19-20]。通过ELISA 检测OrfV 融合基因真核表达质粒pVAX1-B2L-F1L 免疫小鼠血清中Th1(IL-2、IFN-γ)型和Th2 型(IL-4、IL-6)细胞因子含量,结果显示免疫组小鼠经特异性抗原刺激后,分泌产生的IL-2 和IFN-γ细胞因子含量明显升高,且首免后第1 周~第8 周分泌的IL-2和IFN-γ水平均极显著高于本实验室前期研究的pVAX1-B2L、pVAX1-F1L 单独免疫的小鼠;而分泌IL-4 的量与pVAX1-B2L、pVAX1-F1L 单独免疫组相比差异不显著[21];pVAX1-B2L-F1L 组诱导小鼠分泌IL-6 在首免后第2、3、4、6 周均极显著高于pVAX1-B2L 单独免疫组,在首免后第1 周~第5 周极显著高于pVAX1-F1L 单独免疫组;小鼠总体Th1 型细胞因子分泌量大于Th2 型细胞因子分泌量,该结果与赵魁的研究结果一致,表明构建的OrfV pVAXB2L、pVAX-F1L、pVAX-B2L-F1L 质粒主要诱导小鼠产生Th1 型细胞免疫应答。
IL-2 在动物机体的抗病毒、抗毒素、抗肿瘤等方面发挥重要作用[22]。许多研究表明IL-2 作为佐剂在增强疫苗免疫效果的同时,还可以促进机体抗体分泌,缓解临床症状[23]。本研究将pVAX1-IL-2 作为细胞因子免疫佐剂,联合pVAX1-B2L-F1L 免疫小鼠,首免后1~8 周其产生OrfV 特异性抗体水平和脾淋巴细胞增殖水平均高于pVAX1-B2L-F1L 免疫组,且前者小鼠血清中分泌的IL-2和IFN-γ细胞因子含量均高于后者,表明pVAX1-IL-2 能够促进pVAX1-B2L-F1L 诱导小鼠机体中Th1 型细胞因子的分泌;pVAX1-IL-2 对pVAX1-B2L-F1L 诱导小鼠机体分泌IL-4 先有促进作用,后表现为抑制作用,而pVAX1-IL-2 对pVAX1-B2L-F1L 诱导小鼠机体分泌IL-6 无明显促进作用。
本实验研究的OrfV 融合基因真核表达质粒pVAX1-B2L-F1L 免疫小鼠与本实验室前期研究的pVAX1-B2L、pVAX1-F1L 单独免疫小鼠相比,在诱导脾淋巴细胞增殖和Th1、Th2 型细胞因子分泌试验中,前者的免疫效果明显好于单基因真核表达质粒,表明构建的OrfV 双基因融合真核表达质粒比单基因真核表达质量更能增强机体的免疫效果。
