不同生育时期低温胁迫对水稻保护酶的影响
2020-05-30徐伟豪柳洪良朴雪梅韩云哲王亮张基德具红光
徐伟豪 柳洪良 朴雪梅 韩云哲 王亮 张基德 具红光
【摘 要】 通过吉林省内外优质水稻品种为材料,分别在苗期、分蘖期、减数分裂期、灌浆成熟期进行低温胁迫处理,并进行其生理特性研究。结果表明,低温胁迫下水稻的CAT、POD、SOD酶活性均有所增高,其中部分品种的SOD酶活性整体较强,在各个时期均有所增幅,而CAT和 POD酶活性在不同时期存在品种差异,且分蘖期大部分品种的三种保护酶均有明显提升。本试验材料中,不同时期低温胁迫下,通系935、龙稻18、吉粳816、绥粳14、延粳22、通35的整体酶活性在各个时期表现优异,优于其余品种。
【关键词】 水稻;不同时期;低温;耐冷;酶活性
中图分类号:S511 文獻识别码:A 文章编号:2096-1073(2020)05-0049-53
[Abstract] Using high-quality rice varieties inside and outside Jilin Province as materials, they were treated with low temperature stress at the seedling stage, tillering stage, meiosis stage, and filling stage, and their physiological characteristics were studied.The results showed that the CAT, POD, and SOD enzyme activities of rice all increased under low temperature stress. The SOD enzyme activities of some varieties were relatively strong overall, and they increased in all periods, and the CAT and POD enzyme activities were different in different periods, and the three protective enzymes of most varieties at the tillering stage were significantly improved.In this test material, under different periods of low temperature stress, the overall enzyme activity of Tongxi 935, Longdao 18, Jijing 816, Suijing 14, Yanjing 22, and Tong 35 showed excellent performance in each period and was superior to other varieties.
[Key words] rice; different periods; low temperature; cold tolerance; enzyme activity
低温冷害引起水稻减产是世界范围内普遍存在的问题,分布在不同地区的水稻品种在其生育期内常常受到不同程度、不同类型的低温危害[1]。我国东北地区是重要的水稻产区,是我国北方粳稻的主产区,而东北地区的低温冷害发生较为严重[2]。如果在各个生育时期受到低温冷害,会抑制生长发育从而影响其品质和产量[3-5]。
水稻在受到低温冷害之后,植株体内活性氧(ROS)自由基含量明显增加,如果植株体内的抗氧化酶活性受到抑制,过剩的自由基将无法及时得到清除,氧自由基就会在细胞内大量积累,对水稻产生严重伤害[6]。而植株体内存在有效清除ROS的抗氧化酶保护系统,可通过其含量或活性的增加来保护细胞膜免于ROS的伤害,使水稻在一定程度上减缓或抵御逆境伤害[7]。其中, 可以调控ROS代谢平衡的是超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)这3种最主要的酶类[8-11]。
本研究通过引进吉林省和黑龙江省的优质水稻品种,在不同生育时期进行低温胁迫处理,测定其生理指标的变化,筛选出适宜东北地区栽种的耐冷水稻品种,为鉴选高产优质品种提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为从吉林省和黑龙江省引进的不同生育期优质水稻品种共10个,均由选育单位提供(表1)。
1.2 试验方法
1.2.1 耐冷试验 苗期试验将催芽后的种子播种于装有苗床土的育苗盘中,每份材料播15粒,以2cm距离点播,行距3cm,3次重复,在20℃~30℃的温室育苗。在3.5叶龄期,移至人工气候箱进行5℃低温胁迫处理,光照模仿自然低温阴雨天,每天光照12h、黑暗12h,处理7d,以智能温室条件下种植水稻为对照。
分蘖期、减数分裂期、灌浆结实期试验按照常规盘育苗,待3.5~4.0叶移栽至盆内,盆高40cm,直径38cm,每盆栽种4穴,每穴3株基本苗,3次重复。氮、磷、钾纯量分别按120kg/hm2、80kg/hm2、80kg/hm2施用。分别在分蘖期、减数分裂期、灌浆成熟期进行12℃ 6d、16℃ 5d、14℃ 10d低温胁迫处理,以智能温室条件下种植水稻为对照,光照同于苗期。
1.2.2 酶活性测定 苗期、分蘖期处理完成后取全部叶片,减数分裂期、灌浆结实期处理完成后取功能叶片,采集样品后迅速放入密封袋,液氮冷冻后保存于-80℃超低温冰箱,用于保护酶的测定。采用紫外分光光度法测定CAT活性,采用愈创木酚法测定POD活性,采用氮蓝四唑法测定SOD活性[12]。
1.3 数据处理
采用Microsoft Excel 2010和DPS V9.01数据处理系统软件进行相关计算及分析。
2 结果与分析
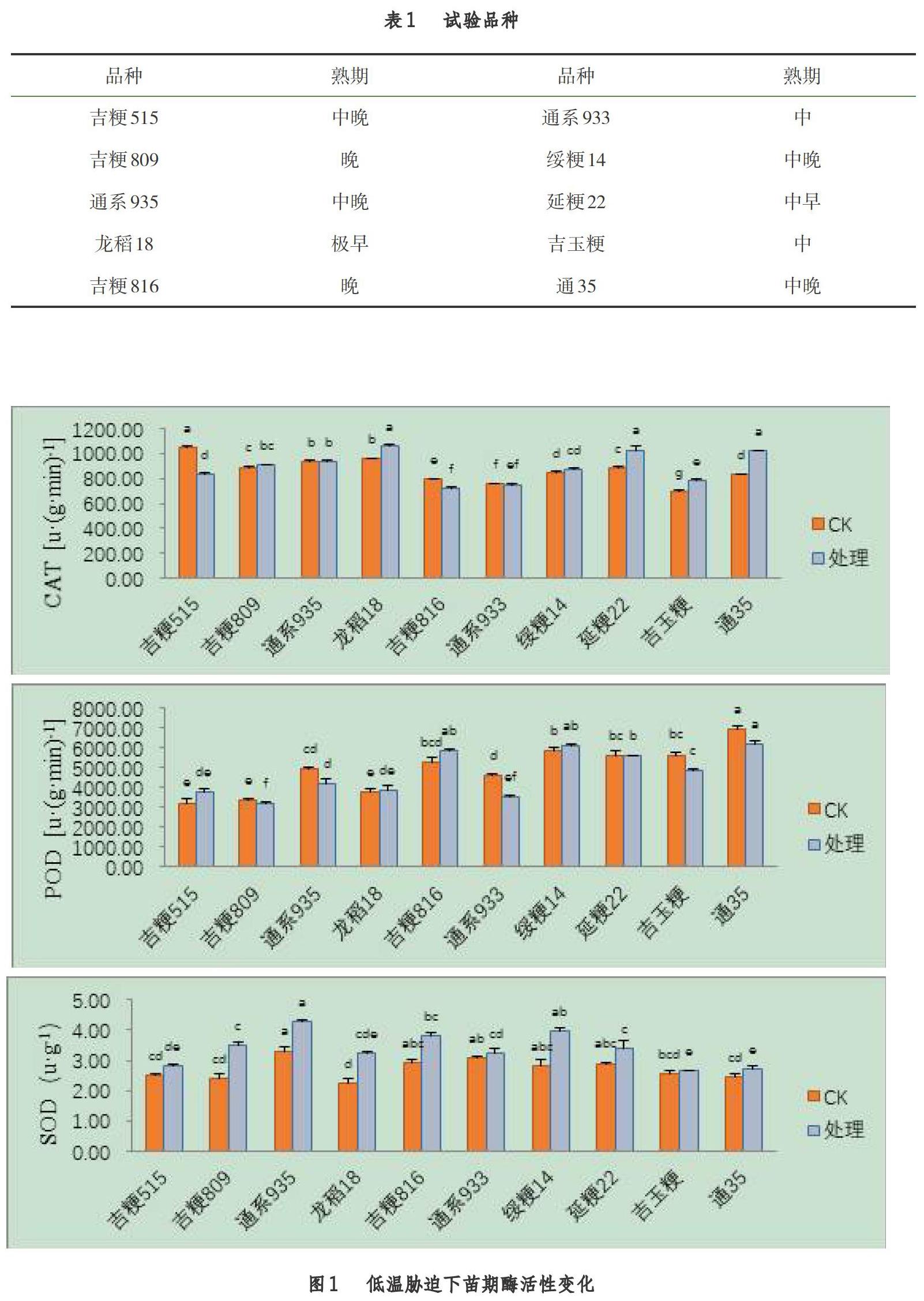
2.1 苗期低温胁迫后对酶活性的影响
由图1可知,不同品种在苗期受低温胁迫后,处理与对照间的酶活性存在显著差异(P<0.05),吉粳809、龙稻18、绥粳14、延粳22、吉玉粳、通35的CAT活性较对照有提升。吉粳515、龙稻18、吉粳816、绥粳14的POD活性较对照有提升。SOD活性是全部品种较对照均有提升。处理品种间来看,不同品种酶活性存在显著差异(P<0.05),龙稻18、延粳22、通35的CAT活性显著高于其余品种,吉粳816、绥粳14、延粳22、通35的POD活性显著高于其余品种,通系935、吉粳816、綏粳14的SOD活性显著高于其余品种。
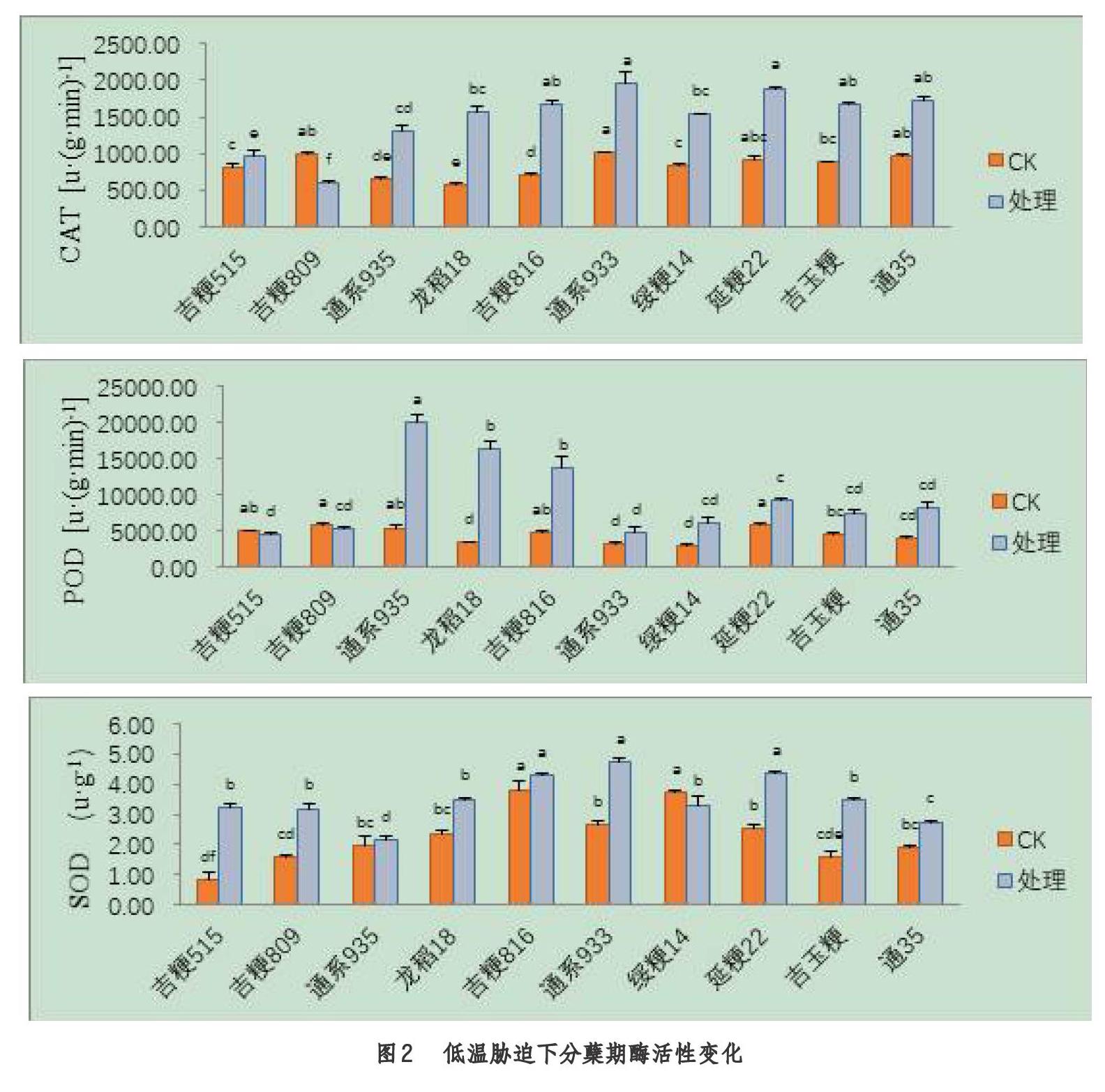
2.2 分蘖期低温胁迫后对酶活性的影响
由图2可知,不同品种在分蘖期受低温胁迫后,处理与对照间的酶活性存在显著差异(P<0.05),大部分品种的酶活性较对照均有提升,且增幅较高。处理品种间来看,吉粳816、通系933、延粳22、吉玉粳、通35的CAT活性显著高于其余品种,通系935、龙稻18、吉粳816的POD活性显著高于其余品种,吉粳816、通系933、延粳22的SOD活性显著高于其余品种。
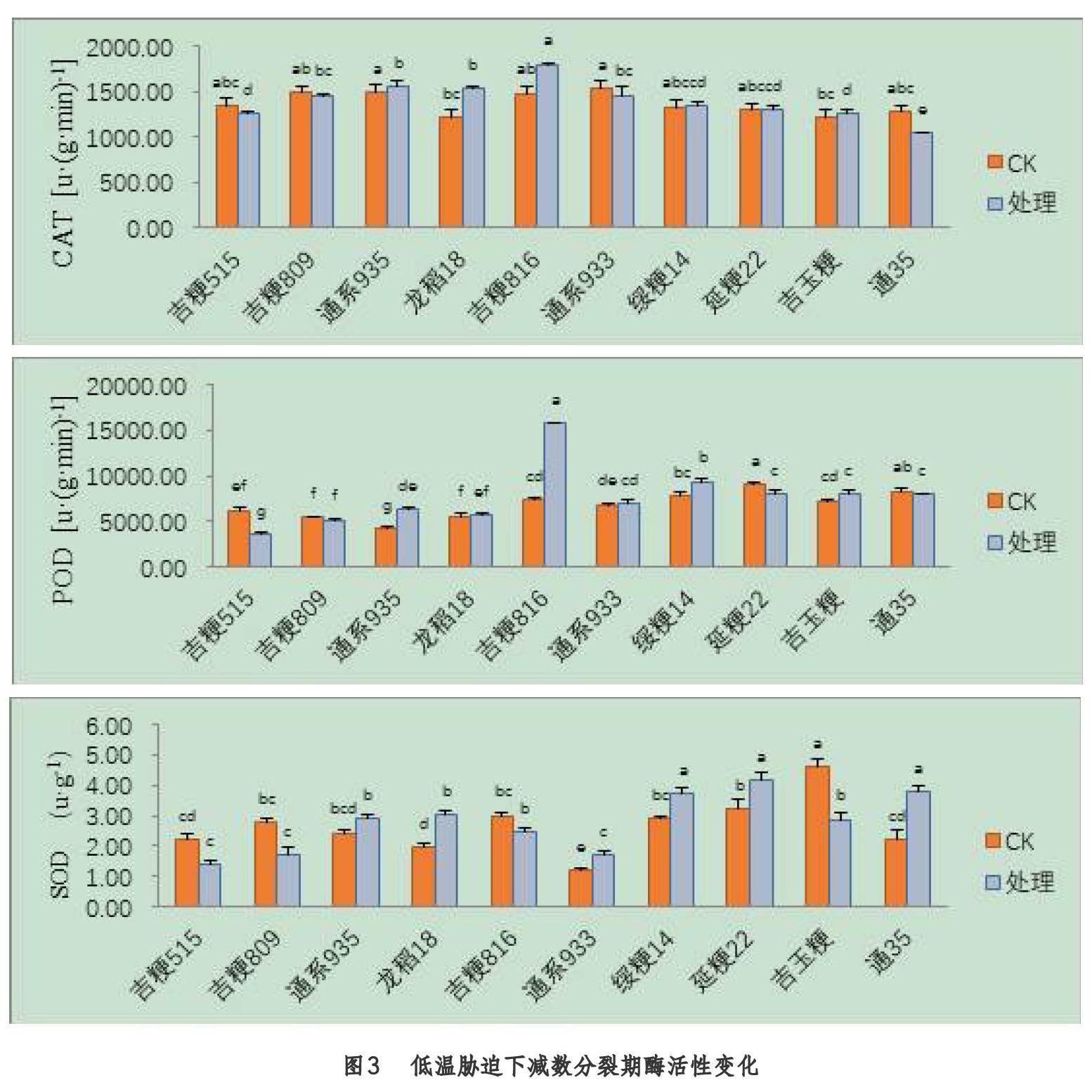
2.3 减数分裂期低温胁迫后对酶活性的影响
由图3可知,不同品种在减数分裂期受低温胁迫后,处理与对照间的酶活性存在显著差异(P<0.05),通系935、龙稻18、吉粳816、绥粳14、吉玉粳的CAT活性较对照有提升。通系935、吉粳816、通系933、绥粳14、吉玉粳的POD活性较对照有提升。通系935、龙稻18、通系933、绥粳14、延粳22、通35的SOD酶活性较对照有提升。处理品种间来看,通系935、龙稻18、吉粳816的CAT活性显著高于其余品种。吉粳816的POD活性显著高于其余品种。绥粳14、延粳22、通35的SOD活性显著高于其余的品种。
2.4 灌浆成熟期低温胁迫后对酶活性的影响
由图4可知,不同品种灌浆成熟期受低温胁迫处理后,处理与对照间的酶活性存在显著差异(P<0.05),吉粳515、吉粳816、绥粳14的CAT活性较对照有提升。吉粳809、通系935、吉粳816、通系933、延粳22的POD活性较对照有提升。吉粳515、吉粳809、吉粳816、绥粳14、延粳22、吉玉粳、通35的SOD活性较对照有提升。处理品种间来看,CAT活性差异不大,其中吉粳816、吉玉粳活性最高。吉粳816、绥粳14、延粳22、吉玉粳的POD活性显著高于其余品种。吉粳809、吉粳816、延粳22的SOD活性较高于其余品种。
3 讨论与结论
本研究结果表明,不同时期大部分品种的SOD酶活性整体较强,在各个时期均有所提升,而CAT和 POD酶活性在不同时期存在品种差异,有的品种是CAT和SOD酶活性较强,POD酶活性相对较弱,有的品种是POD和SOD酶活性较强,CAT酶活性较弱,有的品种是三种酶活性都有较强表现。这说明当处于逆境时,有的品种是三种保护酶相互作用来抵御逆境生长,有的品种则靠一种或两种保护酶的提升来抵御,三种保护酶在植株体内相互协作,共同起到保护作用,且每种酶发挥的作用各不相同,与前人试验结果相类似[13]。本研究还发现在分蘖期大部分品种的三种保护酶均有明显提升,说明分蘖期植株对低温比较敏感,三种保护酶相互协调,共同来抵御逆境进而增强植物的耐冷性。
苗期受低温胁迫后通系935、龙稻18、吉粳816、绥粳14、延粳22、通35酶活性及增幅较高,分蘖期受低温胁迫后通系935、龙稻18、吉粳816、通系933、延粳22酶活性及增幅较高,减数分裂期受低温胁迫后通系935、龙稻18、吉粳816、绥粳14、延粳22、通35酶活性及增幅较高,灌浆成熟期受低温胁迫后吉粳809、吉粳816、绥粳14、延粳22、吉玉粳酶活性及增幅较高。
综合上述,不同时期低温胁迫下,通系935、龙稻18、吉粳816、绥粳14、延粳22、通35的整体酶活性在各个时期表现优异,优于其余品种。
参考文献:
[1] Renata Pereira da Cruz, Raul Antonio Sperotto, Denise Cargn-elutti, et al.Avoiding damage and achieving cold tolerance in rice plants[J].Food and Energy Security, 2013, 2 (2) :96-119.
[2] 侯立刚,马巍,赵国臣,等.吉林省水稻低温冷害发生现状及综合防御措施[J].吉林农业科学,2012,37(04):1-3.
[3] 李婷,张玉霞,王显国,等.低温胁迫下紫花苜蓿根颈抗氧化酶对不同水平磷钾肥处理的响应[J].内蒙古民族大学学报(自然科学版),2017,32(3):243-247.
[4] 杨凤军,李宝江,高玉刚.果树抗寒性的研究进展[J].黑龙江八一农垦大学学报,2003,4:23-29.
[5] 沈祥军,张玉霞,王显国,等.不同水平磷钾肥对沙地紫花苜蓿根颈抗寒性物质的影响[J].内蒙古民族大学学报(自然科学版),2017,32(2):116-121.
[6] 项洪涛,王立志,王彤彤,等.孕穗期低温胁迫对水稻结实率及叶片生理特性的影响[J].农学学报,2016,6(11):1-7.
[7] 于景金,范宁丽,李冉,等.高浓度CO2对热胁迫条件下高羊茅生长和抗氧化系统的影响[J].草业学报,2017,26(8):113-122.
[8] 李晓靖,崔海军.植物对环境胁迫的生理响应研究进展[J].安徽农学通报,2018,24 (14):17-18.
[9] 钱永强,孙振元,韩蕾,等.野牛草叶片活性氧及其清除系统对水分胁迫的响应[J].生态学报,2010,30(7):1920-1926.
[10] 郑殿峰,王广军,宫占元,等.不同播期冬小麦抗寒力的鉴定[J].黑龙江八一农垦大学学报,2000,2:13-16.
[11] 张自阳,冯素伟,姜小苓,等.河南省主推小麦品种越冬期生理生化特性比较研究[J].河南科技学院学报(自然科学版),2012,40 (4):1-6.
[12] 王学奎主编.植物生理生化实验原理和技术[M].北京:北京高等教育出版社,2006.
[13] 陈海燕. 外源硅对低温胁迫下苗期水稻生理生化特性的影响[D]. 哈尔滨: 东北农业大学, 2018.
(编辑:赫亮)
