紫苏Pfgpat9基因的鉴定与功能分析
2020-05-28任文燕郝月茹李润植周雅莉杨宏斌王计平
任文燕,郝月茹,李润植,周雅莉,杨宏斌,王计平
(山西农业大学农学院, 太谷 030801)
紫苏[Perillafrutescens(L.)Britt.],又名香苏、白苏、赤苏,为一年生自花授粉作物,是国家卫生部颁布的首批兼保健药用、食用的60种植物之一[1-4],在中国种植已经有2 000多年的历史。紫苏种子富含油脂,含油率可达46%~55%,油脂中含多种脂肪酸,主要包括亚麻酸(C18∶3)、油酸(C18∶1)、亚油酸(C18∶2)等不饱和脂肪酸[5],其中人体必需脂肪酸α-亚麻酸含量高达55%~65%,比大豆等油料作物α-亚麻酸含量高出5倍左右[6]。
甘油三磷酸酰基转移酶(glyceryl triphosphate acyltransferase,GPAT)是三酰甘油(triacylglycerol,TAG)生物合成的限速酶[7,8],催化Kennedy途径的第一步反应,主要将酰基从acyl-ACP或acyl-CoA转移到甘油-3-磷酸(glycerol-3-phosphate,G3P)的sn-1位置上形成溶血磷脂酸(lysophosphatidic acid,LPA)[9-11]。目前已经在拟南芥中鉴定出10个gpat基因[12-14],已有研究表明,拟南芥gpat9(Atgpat9)在TAG的生物合成中起直接作用[12,15]。Shockey等[16]研究表明,使用人工合成的miRNA使Atgpat9基因沉默,降低了拟南芥种子油含量。Singer等[17]发现Atgpat9在拟南芥发育叶片的极性脂质、TAG的生物合成以及花粉粒脂滴的合成中起重要作用。Jain等[18]将克隆到的红花质体gpat基因转化至拟南芥,转基因拟南芥种子含油量比野生型增加了10%~21%。Miriam等[19]在酵母互补试验中表达向日葵gpat9(Hagpat9) 基因,使酵母TAG总量增加,并且脂肪酸组分也发生了改变,表现在C16∶0和C16∶1脂肪酸的富集。目前,已经在拟南芥(Arabidopsisthaliana)[20]、甘蓝型油菜(Brassicanapus)[21]、向日葵(Helianthusannuus)[22]等多种植物中克隆到gpat基因,并且异源表达该基因可以提高宿主组织的油脂含量。而关于紫苏gpat9(Pfgpat9)基因的研究还未见报道。
本研究基于前期紫苏转录组数据分析中获得的高表达gpat基因的转录本,以‘晋紫苏1号’发育前期(开花后20 d)的种子为试验材料,利用RT-PCR技术克隆得到Pfgpat9基因完整的开放阅读框(open reading frame,ORF)序列,并分析该基因在不同器官组织的表达特性。构建酵母表达载体,转化至野生型酵母INVSc1和缺陷型酵母gat1,分析转基因酵母脂肪酸组分及含量,鉴定Pfgpat9基因的生物学功能。该研究为深入解析紫苏油脂合成分子机制提供理论依据。
1 材料与方法
1.1 供试材料
试验所用紫苏品种为‘晋紫苏1号’,2018年4月种植于山西农业大学实验基地,对紫苏根、茎、叶、花及四个不同发育时期的种子(即开花后10、20、30、40 d)分别取样,液氮速冻后置于-80 ℃冰箱保存备用。大肠杆菌感受态DH5α、试验所用克隆载体pMD18-T、表达载体pYES2.0、酿酒酵母野生型菌株INVSc1和缺陷型菌株gat1均保存于山西农业大学分子农业与生物能源研究所。
1.2 试验方法
1.2.1 紫苏总RNA提取及cDNA合成
采用EASY spin Plus RNA 快速提取试剂盒(RN09,北京艾德莱生物科技有限公司)提取紫苏RNA,反转录参考ABM公司5X All-In-One MasterMix(with AccuRT Genomic DNA Removal Kit)反转录试剂盒完成。
1.2.2Pfgpat9基因的克隆
本试验从紫苏转录组数据库进行筛选比对,获得高表达的编码甘油三磷酸酰基转移酶9转录本,命名为Pfgpat9。以该转录本序列为模板,设计PCR扩增引物(表1),按照全式金2×EasyTaq®PCR SuperMix说明书的反应体系与反应程序;以紫苏开花后20 d的种子cDNA为模板,进行PCR扩增;割胶回收PCR产物后,将其克隆至pMD18-T载体上,得到pMD18-T-Pfgpat9并转化大肠杆菌DH5α;菌液PCR鉴定筛选阳性克隆,送通用生物公司(安徽)进行测序鉴定。
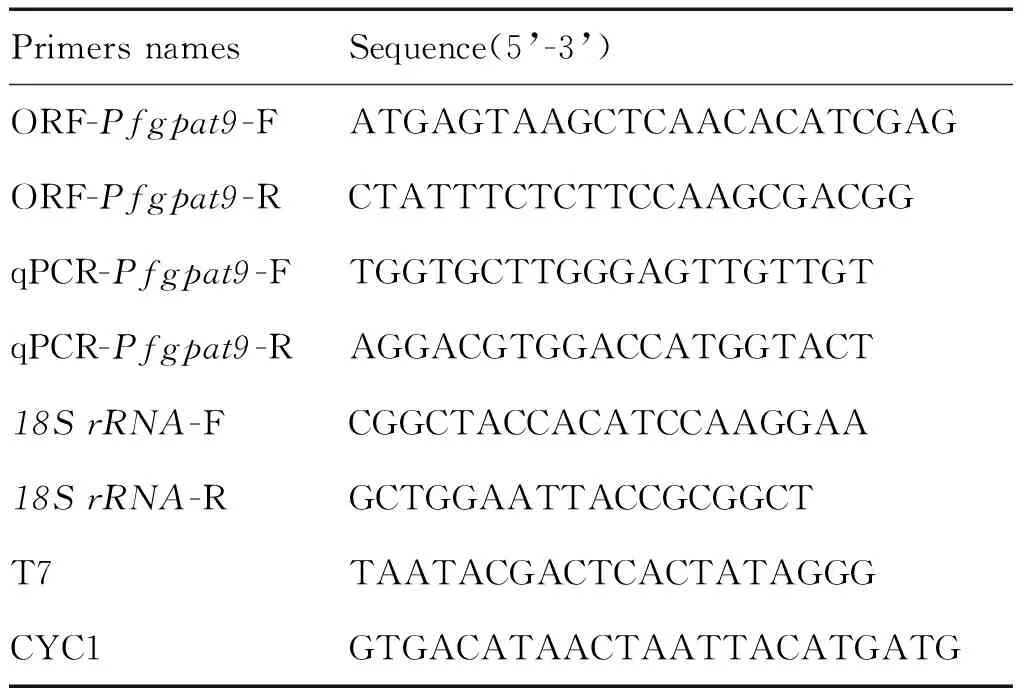
表1 引物信息Tab.1 Primer information used in this study
1.3 Pfgpat9基因的生物信息学分析
使用在线软件ProParam对PfGPAT9蛋白的理论等电点、分子量、不稳定性等进行分析。采用软件TMHMM对PfGPAT9的跨膜结构域进行预测。通过在线软件SOPMA和SWISS-MODEL对PfGPAT9蛋白进行二级结构及三级结构的预测和建模。利用NCBI-CDD预测PfGPAT9蛋白的功能结构域。采用在线软件PRALINE program对紫苏PfGPAT9与其他物种的氨基酸序列进行比对。运用MEGA7.0对紫苏和其他物种的GPAT9蛋白构建系统发育树。
1.4 实时荧光定量PCR检测
实时荧光定量PCR使用ABM公司的EvaGreen 2× qPCR试剂盒,并设计qPCR引物(表1),以18S rRNA作为内参基因[23],进行PCR检测。反应体系(10 μL)为:cDNA模板0.5 μL,正反向引物各0.3 μL,EvaGreen 2× qPCRMaster Mix 5 μL,Nuclease-free H2O 3.9 μL,设置3次生物学重复。反应程序采用两步法进行:95 ℃ 10 min,95 ℃ 15 s,58 ℃ 1 min,40个循环。采用2-△△Ct法计算紫苏Pfgpat9基因的相对表达量[24]。所得数据通过SPSS软件进行统计分析。
1.5 Pfgpat9基因酵母表达载体构建及转化
本试验所用酵母表达载体为pYES2.0,含URA3位点和氨苄青霉素抗性。Pfgpat9基因通过EcoR I和NotI双酶切连接到pYES2.0中形成重组酵母表达载体pYES2.0-Pfgpat9,将其转入大肠杆菌DH5α感受态细胞中,进行菌液PCR鉴定,提取质粒进行双酶切验证。
将鉴定为阳性的重组质粒和空载体分别转化至野生型酵母菌株INVSc1和缺陷型菌株gat1,28 ℃培养2~4 d后通过菌液PCR鉴定阳性克隆菌株,用于后续试验。菌液PCR体系(20 μL)为:菌液0.4 μL,2×San Taq PCR Mix 10 μL,10 μmol/L正反引物(Pfgpat9基因检测引物为Pfgpat9-F和Pfgpat9-R,空载对照质粒检测引物为T7和CYC1)各0.8 μL、dd H2O 8 μL。扩增程序为:94 ℃预变性5 min;94 ℃变性30 s,58.5 ℃退火30 s,72 ℃延伸90 s,循环30次;72 ℃终延伸10 min。
培养酵母细胞的方法参考徐荣华[25]、谭太龙[26]等,将转Pfgpat9基因的酵母与转空载体对照质粒的酵母在SC-U(synthetic minimal defined medium lacking uracil)抑制培养基(2%葡萄糖)中培养,然后转至SC-U诱导培养基(2%半乳糖)中继续培养,诱导表达36 h后离心收集菌体,冷冻干燥后进行脂肪酸含量与组分分析。
1.6 转基因酵母的总脂肪酸含量测定及脂肪酸组分分析
总脂肪酸的提取及测定:试验采用甲醇与氯仿抽提的方法来提取总脂。称取50 mg转基因酵母菌体粉末置于离心管中,加入甲醇与氯仿(体积比为2∶1)混合液7.5 mL,37 ℃提取24 h;再次加入甲醇与氯仿混合液7.5 mL,提取12 h,抽提2次。将三次所得有机相混合,再加入一定比例1% NaCl溶液和氯仿溶液,离心吸取下层有机相至已经称重的新离心管m0,氮吹仪吹干后称管重m1,每个样品3次重复。总脂肪酸含量=(氮吹仪吹干后的管重m1-加样前的管重m0)/酵母菌体粉末重量。
脂肪酸的提取与组分分析参考王计平等[23]分析方法。试验使用Tri 17∶0作为内标,用2.5%甲醇(浓硫酸)进行甲酯化,80 ℃水浴2 h,用0.4 mL正己烷溶解脂肪酸,转至GC瓶用于气相检测。用安捷伦气相色谱仪7890B对脂肪酸各组分含量进行分析。
2 结果与分析
2.1 Pfgpat9基因的克隆
以紫苏开花后20 d的种子cDNA作为模板,进行PCR扩增,割胶回收目的片段,纯化测序,获得一条1 116 bp的核苷酸序列,并且测序结果与转录组数据比对分析结果一致,确定该片段为Pfgpat9基因序列(图1)。
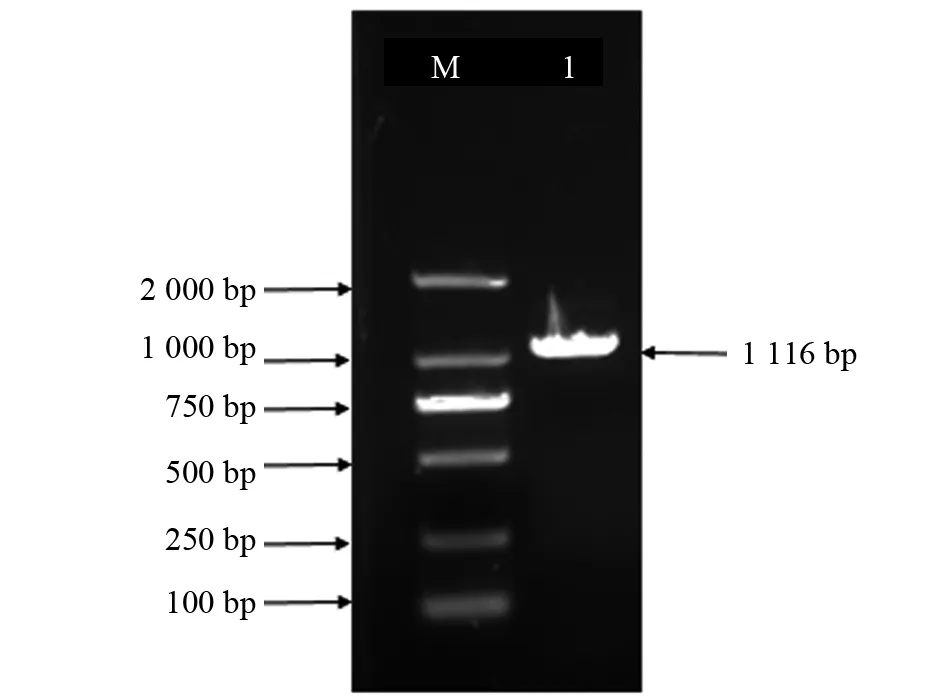
图1 Pfgpat9基因PCR扩增产物电泳结果Fig.1 Electrophoresis of the amplified PCR product of Pfgpat9 M:DL2000 Marker;1:Pfgpat9基因扩增产物M:DL2000 Marker;1:The PCR product of Pfgpat9
2.2 Pfgpat9基因编码蛋白的序列特征
2.2.1PfGPAT9蛋白的理化性质
对Pfgpat9基因的序列结构分析表明(图2),Pfgpat9基因的ORF为1 116 bp,共编码371个氨基酸,蛋白相对分子量为42.92 kD,理论等电点(pI)为8.96,不稳定系数为46.67,平均亲水系数为-0.108,脂质指数为93.45,因此推导该蛋白质为亲水性不稳定蛋白。跨膜结构域结果分析显示:PfGPAT9蛋白分别在第69~91位、95~117位和130~147位氨基酸之间存在3个典型的跨膜螺旋区。

图2 Pfgpat9 ORF序列及编码的氨基酸序列Fig.2 Sequence of Pfgpat9 ORF and the translated amino acid sequence
运用SOPMA软件对PfGPAT9蛋白进行二级结构预测,结果显示:该蛋白的二级结构主要包括α-螺旋(40.16%)、无规则卷曲(31.81%)、延伸链(21.02%)、β-折叠(7.01%)。其中,α-螺旋和无规则卷曲是PfGPAT9蛋白的主要结构元件。
利用SWISS-MODEL在线软件以结核分枝杆菌(Mycobacteriumtuberculosis)GPAT蛋白[27]为模板(图3b),对PfGPAT9蛋白进行同源建模,结果发现,PfGPAT9蛋白与模板蛋白的序列相似性为64%,氨基酸保守序列范围为104~347位氨基酸,表明建立的模型可信度高。对比图3a和图3b可以发现,PfGPAT9蛋白与模板蛋白的外形结构高度相似,可以推测PfGPAT9蛋白与模板蛋白有着相同的亚基BMA(beta-D-mannose)(图3c)和PLM(palmitic acid)(图3d)。NCBI-CDD分析该蛋白的功能结构域,发现PfGPAT9蛋白含有PLN02833保守结构域,属于溶血磷脂酰基转移酶(lysophospholipid acyltransferase superfamily,LPLAT)超家族中的成员。
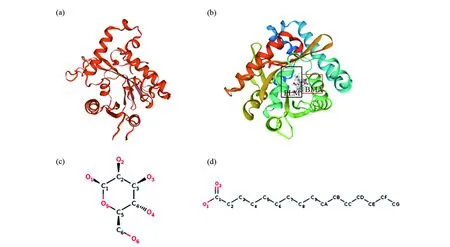
图3 PfGPAT9蛋白三级结构预测Fig.3 The tertiary structure prediction of GPAT9 protein in Perilla frutescens (a)PfGPAT9蛋白的三维结构;(b)结核分枝杆菌蛋白的三维结构;(c)配体BMA;(d)配体PLM(a)Tertiary structure of PfGPAT9 protein of Perilla frutescens;(b)Tertiary structure of Mycobacterium tuberculosis;(c)Ligand BMA;(d)Ligand PLM
2.2.2PfGPAT9蛋白多序列比对与系统发育分析
通过PRALINE program对紫苏、拟南芥(Arabidopsisthaliana)、大豆(Glycinemax)、甘蓝型油菜(Brassicanapus)、油桐(Verniciafordii)等植物的氨基酸序列进行比对,并用MEGA7.0构建系统进化树。序列比对分析表明(图4):GPAT9蛋白的分化程度较低,氨基酸序列高度保守,相似度高达90%以上;GPAT9蛋白具有典型的酰基转移酶结构域,并且包含4个高度保守的基序。系统发育树结果显示(图5),紫苏与油橄榄(Oleaeuropaea)的亲缘关系最近。
2.3 Pfgpat9基因表达特性分析
选取紫苏根、茎、叶、花及四个不同发育时期的种子为材料,对Pfgpat9基因进行表达特性分析。结果表明(图6):该基因在紫苏根、茎、叶、花及种子中均有表达,其中在种子中的表达量明显高于其他组织,并且在种子不同发育时期,该基因的表达量存在显著差异,在开花后20 d表达量达到最高,为开花后10 d的2.47倍。
2.4 酵母表达载体的构建
利用EcoR I和NotI对重组质粒pYES2.0-Pfgpat9进行双酶切,以空载体作为对照。图7为双酶切后的电泳图,其中第一泳道为双酶切后得到的目的基因条带,其长度大小为1 116 bp,与目的基因Pfgpat9的长度大小一致,第二泳道为空载体(5.9 kb)。这说明酵母表达载体pYES2.0-Pfgpat9构建成功,可用于后续试验。
2.5 转基因酵母的阳性鉴定
将构建好的酵母表达载体转入野生型酵母INVSc1和缺陷型酵母gat1中,使用SC-U诱导培养基(2%半乳糖)进行筛选,挑取单克隆菌落培养后进行阳性检测。检测结果见图8,均扩增出目的条带,表明目的基因已成功转入酵母中。
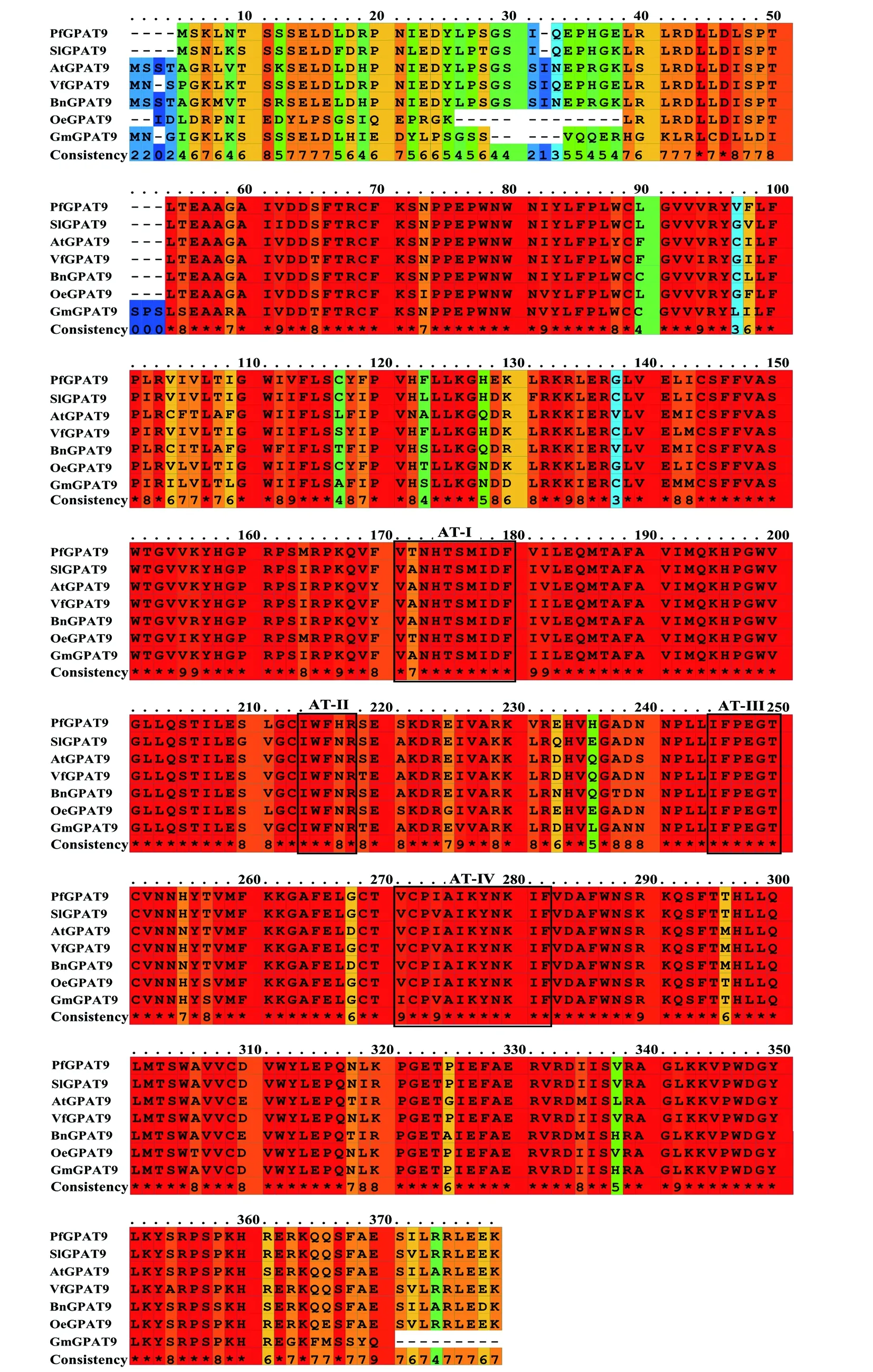
图4 不同物种的GPAT9氨基酸序列比对分析(黑色方框代表GPAT9保守结构域)Fig.4 Sequence alignment of GPAT9 proteins from different plant species (Black box represents the GPAT9 conserved motifs)
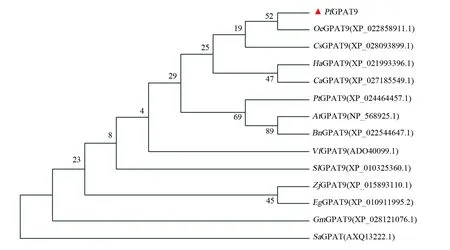
图5 PfGPAT9与其他物种GPAT9蛋白的系统发育分析Fig.5 Phylogenetic tree of GPAT9 proteins of Perilla frutescens and other species Oe:油橄榄;Cs:茶树;Ha:向日葵;Ca:小果咖啡;Pt:黑色杨木;At:拟南芥;Bn:甘蓝型油菜;Vf:油桐;Sl:番茄;Zj:枣;Eg:非洲油棕;Gm:大豆; Sa:海藻施万氏菌Oe:Olea europaea;Cs:Camellia sinensis;Ha:Helianthus annuus;Ca:Coffea arabica;Pt:Populus trichocarpa;At:Arabidopsis thaliana;Bn:Brassica napus;Vf:Vernicia fordii;Sl:Solanum lycopersicum;Zj:Ziziphus jujuba;Eg:Elaeis guineensis;Gm:Glycine max;Sa:Shewanella algae
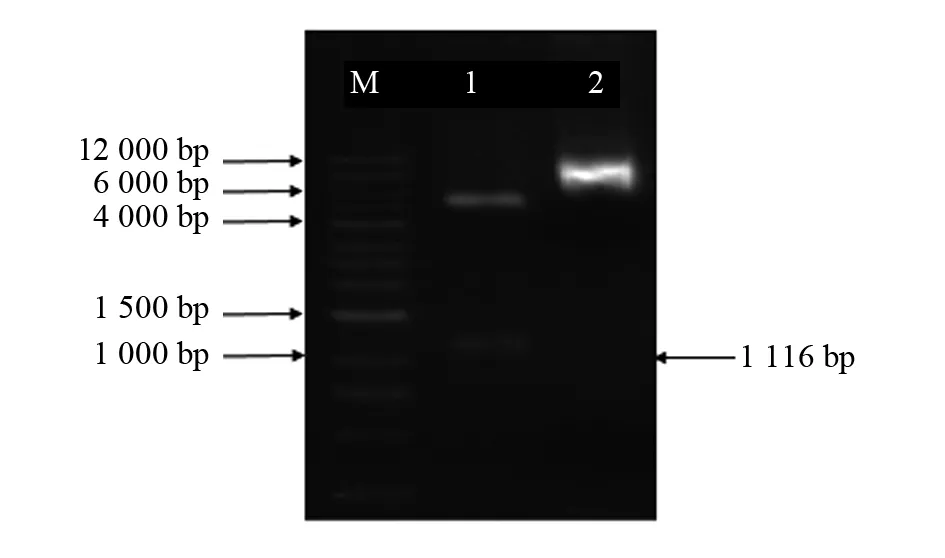
图7 酵母表达载体双酶切鉴定Fig.7 Double digestion of the yeast expression vectorM:DL12000 Marker;1:pYES2.0-Pfgpat9双酶切结果;2:空载体pYES2.0M:DL12000 Marker;1:Double digested electrophoresis of pYES2.0-Pfgpat9;2:pYES2.0 vector
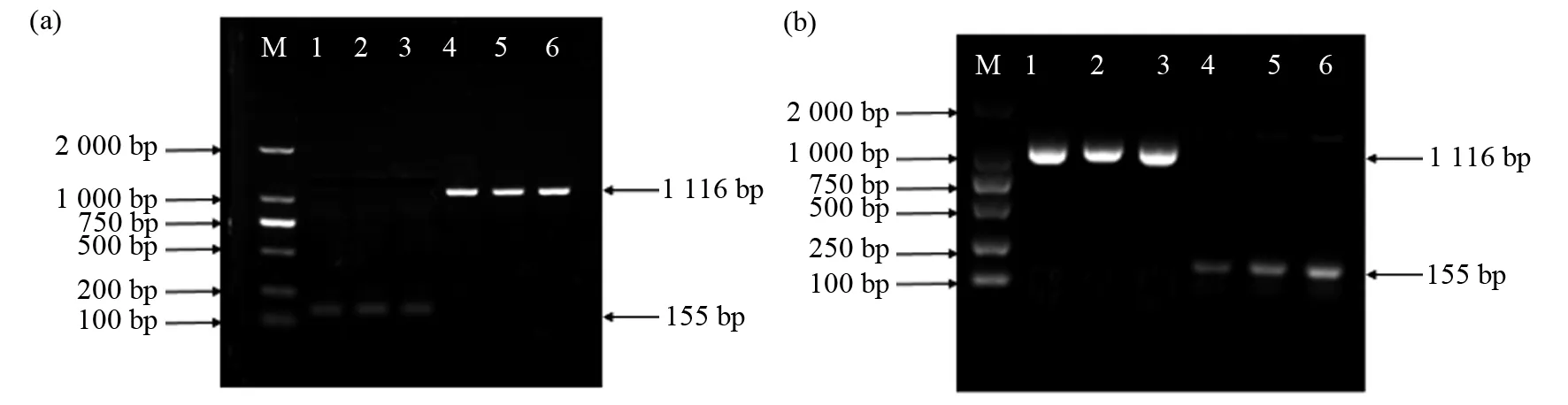
图8 转Pfgpat9菌液PCR产物琼脂糖凝胶电泳Fig.8 Detection of Pfgpat9 colony PCR products were analyzed on agarose gels(a)M:DL2000 Marker;1~3:INVSc1转空载对照质粒pYES2.0;4~6:INVSC1转pYES2.0-Pfgpat9;(b)M:DL2000 Marker;1~3:gat1转pYES2.0-Pfgpat9;4~6:gat1转空载对照质粒pYES2.0(a)M:DL2000 Marker;1~3:INVSc1 transformed with empty pYES2.0;4~6:INVSc1 transformed with pYES2.0-Pfgpat9;(b)M:DL2000 Marker;1~3:gat1 transformed with pYES2.0-Pfgpat9;4~6:gat1 transformed with empty pYES2.0
2.6 转基因酵母脂肪酸含量及组分分析
选择野生型INVSc1、缺陷型gat1、转野生型空载体、gat1-pYES2.0空载体、转pYES2.0-Pfgpat9野生型酵母、转pYES2.0-Pfgpat9缺陷型酵母菌株进行总脂肪酸含量、脂肪酸组分及含量的测定。结果显示,转基因INVSc1酵母总脂肪酸含量比转空载酵母(empty vector,EV)提高了2.2%,比野生型酵母总脂含量高1.01%(图9a),转基因gat1酵母总脂肪酸含量比EV提高了1.55%,比缺陷型酵母总脂含量高1.68%(图9b)。各脂肪酸组分及含量分析表明,转基因INVSc1酵母与EV相比C16∶0、C16∶1、C18∶1含量均有所提高,分别提高了14.57 %、15.28%和1.12%,C18∶0含量减少0.84%,并且转基因酵母新合成了C18∶2,其含量为7.28%,与野生型酵母相比C16∶0、C16∶1、C18∶1分别提高了14.44%、16.42%、1.5%,C18∶0含量减少0.99%(图10a)。转基因gat1酵母与EV相比C16∶0、C16∶1、C18∶0均有所提高,分别提高7.33%、23.52%和8.22%,C18∶1含量减少1.62%,与缺陷型酵母相比C16∶0、C16∶1、C18∶0、C18∶1均提高,分别提高了12.55%、19.74%、8.34%和2.18%(图10b)。转基因酵母油脂成分的变化充分证明了该基因参与酵母细胞的油脂合成代谢。
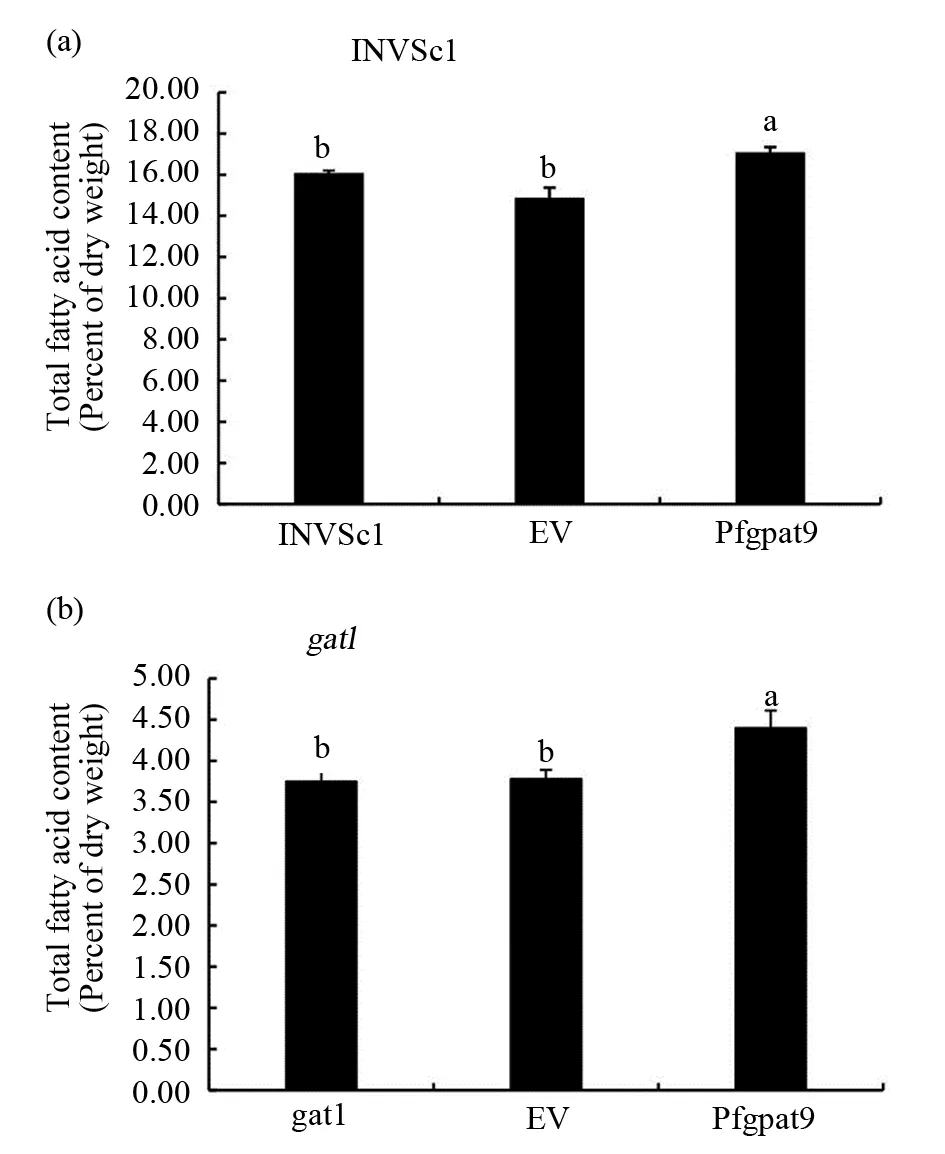
图9 转基因酵母总脂肪酸含量Fig.9 Total lipids content in the transgenic yeast(a)野生型酵母总脂肪酸含量;(b)缺陷型酵母总脂肪酸含量(a)Total lipids content in the INVSc1;(b)Total lipids content in the gat1
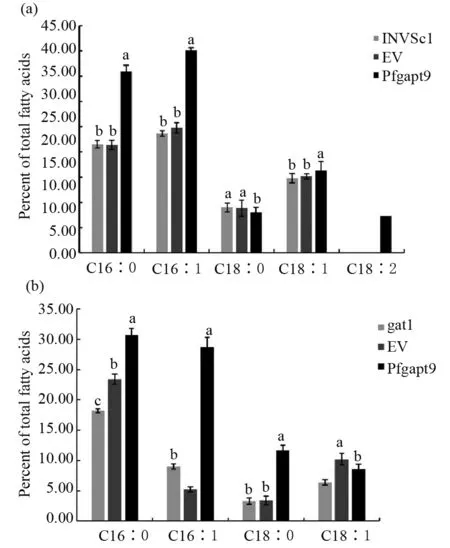
图10 转基因酵母的脂肪酸组分分析Fig.10 Composition analysis of fatty acids in transgenic yeasts(a)野生型酵母的脂肪酸组分分析;(b)缺陷型酵母的脂肪酸组分分析(a)Composition analysis of fatty acids in INVSc1; (b)Composition analysis of fatty acids in gat1
3 讨论
GPAT是油料作物油脂合成积累途径中催化三酰甘油生物合成第一步反应的关键酶。目前,已从拟南芥、向日葵、油桐等植物中克隆到gpat9基因,并通过试验证明过表达该基因可以使TAG的含量显著升高[17]。本研究克隆获得紫苏Pfgpat9基因,并对其编码蛋白序列特征进行了分析,结果表明,该基因ORF序列长为1 116 bp,编码371个氨基酸,相对分子量为42.92 kD。对PfGPAT9蛋白特性分析发现,该蛋白为亲水性不稳定蛋白。跨膜结构分析表明,PfGPAT9蛋白存在3个跨膜区,这与AtGPAT9蛋白含有3个跨膜结构域的特征一致[12]。多序列比对结果显示,PfGPAT9蛋白具有酰基转移酶活性结构域,符合GPAT酶蛋白的典型特征,对PfGPAT9蛋白的系统进化分析表明,紫苏PfGPAT9与油橄榄、茶树等亲缘关系较近,而与大豆、油棕的关系较远。
植物种子发育过程中,TAG积累模式为:在发育初期油脂积累缓慢,发育中期TAG快速积累,在种子发育后期TAG趋于饱和,合成量降低[26]。本研究通过qRT-PCR分析发现,Pfgpat9基因在紫苏的各个器官中均有表达,但在种子中的表达量显著高于其它器官,在开花后20 d种子中表达量最高,这与向日葵[19]Hagpat9基因和拟南芥[17]Atgpat9基因表达趋势基本一致。Pfgpat9基因表达模式与种子发育过程中油脂积累模式相一致,表明两者之间呈现正相关,推测Pfgpat9基因在紫苏种子油脂合成积累中起重要作用。
Sui等[28]研究结果表明,番茄gpat基因的过量表达增加了类囊体膜中的不饱和脂肪酸含量;陈文玲等[29]发现gpat9直接参与真核细胞中膜脂和油脂的合成;张楠等[30]在酵母中异源表达小桐子gpat基因,显著提高了酵母的总含油量。为进一步鉴定紫苏Pfgpat9基因的功能,本研究通过转化野生型酵母和缺陷型酵母,对转基因酵母总油脂及其脂肪酸组分进行分析,结果表明,Pfgpat9基因显著提高了酵母总脂肪含量,且引起了酵母脂肪酸组分的变化,在转野生型酵母中新合成了C18∶2,并且C16∶0、C16∶1、C18∶1含量均有所提高,在转缺陷型酵母中C16∶0、C16∶1、C18∶0含量均提高。Miriam等[19]在酵母互补试验中,过表达Hagpat9增加了酵母细胞中C16∶0和C16∶1脂肪酸的含量,这与本试验结果一致。
本文从紫苏发育种子中成功克隆Pfgpat9基因,其编码蛋白具有较高的GPAT酶活性,异源表达结果表明,Pfgpat9基因可显著促进宿主组织总油脂合成以及C16∶1脂肪酸的富集。本研究为深入解析紫苏种子油脂合成机制及用于其他油料作物品质改良提供了新技术和新的基因元件。
