水稻叶龄变化对短日长的响应
2020-05-27王亚梁朱德峰陈惠哲向镜张义凯徐一成张玉屏
王亚梁 朱德峰 陈惠哲 向镜 张义凯 徐一成 张玉屏


摘要:光周期是影响水稻叶片数的重要环境因子。本研究利用盆栽试验研究了10 h短日长处理对水稻品种叶片数的影响。结果表明,对生产中主茎总叶片数14~17叶的水稻品种来说,除3叶期外,短日长处理不同程度减少水稻主茎总叶片数,7~9叶龄期对短日长最敏感,7叶期和9叶期短日长处理使水稻主茎总叶片数显著减少。7叶期短日处理,水稻主茎总叶片数减少幅度表现为:高短日长敏感型品种>中短日长敏感型品种>低短日长敏感型品种。短日长处理对所有品种穗分化期叶龄余数影响不大。在不同品种类型中,籼粳杂交稻对短日长响应最为敏感,短日长处理下籼粳杂交稻主茎叶片数下降幅度大于粳稻和籼稻。在同一亚种内不同品种对短日长响应的敏感性也存在差异。
关键词:水稻;短日长;叶龄;品种差异
中图分类号:S511.01文献标识码:A文章编号:1000-4440(2020)02-0265-06
Abstract:Day length is an important environmental factor affecting the number of rice leaves. In this study, the effects of 10 hours short day treatment on the number of rice leaves were studied by pot experiment. The results showed that, except for the 3-leaf stage, the final main-stem leaf number of rice with14-17 leaves was reduced under short day length treatment. Moreover, the final main-stem leaf number was significantly decreased under the shortday length treatment at 7-leaf stage abd 9-leaf stage. The reduction of final main-stem leaf number after short day length treatment at the 7-leaf stage of rice presented the tendency as follows: high short-day sensitive cultivars > medium short-day sensitive cultivars > low short-day sensitive cultivars. The short-day length did not influence the leaf age remainder at panicle initiation stage of every rice cultivar. For different subspecies, indica-japonica hybrid rice was most sensitive to short day length treatment. In addition, the reduction of final main-stem leaf number of indica-japonica hybrid rice was higher than that of japonica rice and indica rice under short day length treatment. In conclusion, the sensitivity of different varieties in the same subspecies to short day length is different.
Key words:rice;short day length;leaf age;cultivar difference
水稻是中國主要的粮食作物,其种植区域广、品种类型多[1]。在水稻生长的季节,北方稻区日照长,南方稻区日照短。水稻品种生育期受种植地温度和日照长度的影响,同一品种在不同地区和季节种植生育期变化较大,因此,不同地区引种要根据品种的光温反应特性和当地的日长和温度确定。粳稻和籼稻的生态适应性不同,南方稻区水稻品种以籼稻为主,北方稻区水稻品种以粳稻为主。 随中国社会经济发展和生活水平提高,南方居民对粳米需求增长。近年来,南方稻区原种植籼稻的地区引入和推广粳稻、籼粳杂交稻品种,特别是长江中下游稻区的单季粳稻种植面积不断扩大[2-3]。
水稻叶片与茎、根系的生长和穗发育存在以水稻叶片为指标的器官同伸关系[4-5]。水稻栽培中提出叶龄模式,按主茎总叶龄(N)和伸长节间数(n)将品种进行归类,确定有效分蘖临界期(N-n和N-n-1)、拔节叶龄期(n-2的倒数叶龄期)和穗分化叶龄期(倒3.5叶)等关键叶龄期的规律,为水稻肥水管理、群体调控提供叶龄指标。
水稻主茎叶片数在不同水稻种植地区和种植季节差异很大,同一地区水稻播期推移也会减少水稻总叶片数[6-7]。叶龄是品种特性,受温度和日长等环境因素的影响。环境因素对水稻叶龄影响的研究结果表明,温度升高会提高水稻的出叶速率,有效积温和主茎叶片数具有显著的正相关关系 [8-11]。日照长度也对主茎叶片数变化造成影响,主茎叶片数的变化间接体现在水稻生育期上,唐锡华等[12]指出,除少数长日照水稻品种外,大多数水稻品种生育期受日长影响大于温度。研究认为,水稻营养生长期对光周期最为敏感,但也发现光周期在水稻生殖生长期对水稻的生长发育造成影响[13]。基本营养生长期后,水稻开始感受光周期,但同时受到温度的互作影响[14]。水稻品种对光周期的反应存在品种间差异,Yin等[13]发现籼稻的基本营养生长期要比粳稻长,因此表现出较弱的光周期敏感性。孟亚利等[15]报道指出,水稻在4叶期开始感光,但不同水稻进入生殖生长所需经历的短光周期是不一致的[16]。
中国水稻品种不同地区引种及不同季节种植,水稻生长期间日长变化较大,水稻主茎总叶片数发生变化。目前,水稻品种主茎总叶片数受日长影响而变化的敏感时期尚不清楚,品种间差异也不明确。为此,本研究选择有代表性的主导品种,研究水稻品种总叶片数对日长的响应及品种间差异,为不同地区水稻引种、叶龄模式在水稻栽培上的应用提供理论依据。
1材料与方法
1.1供试品种
选用中国主要稻区生产上应用的主导水稻品种25个,品种主要适应区域见表1。
1.2短日照处理
采用盆栽方法,进行水稻短日照处理。水稻短日照处理按叶龄设置,当水稻叶龄分别生长到3叶、5叶、7叶、9叶、11叶、13叶时,将种植盆移入短日照处理装置。短日照处理装置利用不锈钢搭成,长、宽和高分别为10.0 m、3.0 m和1.6 m,利用透气遮光布盖住遮光。短日照处理时间为17∶00至第2 d早上7∶00,每天日照长度为10 h,连续处理7 d。每处理3次重复,每重复10盆。
1.3种植方法
试验在中国水稻研究所试验场进行。利用塑料种植盆种植,种植盆规格为20 cm×18 cm×25 cm。每盆装过筛土10 kg, 土壤理化性质为:pH 5.93、 有机质28.26 g/kg、 全氮1.50 g/kg、 碱解氮128.24 mg/kg、 全磷0.87 g/kg、 有效磷44.8 mg/kg、 全钾25.27 g/kg、 速效钾130.00 mg/kg。于5月21日播种,基质育秧,6月1日选长势一致的秧苗移栽至塑料种植盆中,每盆种4株。基肥每盆施尿素0.5 g,过磷酸钙1.62 g,氯化钾0.26 g,移栽7 d后施尿素0.25 g作分蘖肥,穗分化期施尿素0.50 g和氯化钾0.26 g。水稻生长期间其他管理措施与高产栽培一致。
1.4测定项目与方法
1.4.1叶龄记载移栽后对主茎挂牌,每隔5 d记录主茎叶龄,叶龄记载参照Haun等[17]的方法。
1.4.2穗分化期确定水稻7叶期后每隔3 d,选取主茎3个,剥掉莖鞘后用体视显微镜(OLYMPUS, SZX9)检查幼穗分化,确定穗分化时期。
穗分化期叶龄指数= 穗分化期叶龄/总叶片数×100%。
1.5数据统计与分析
采用Excel统计数据,用SAS 9.2软件进行聚类分析(最长距离法),利用Duncans法(新复极差法,SSR)检验处理间差异。
2结果与分析
2.1水稻主茎总叶片数对短日长处理的敏感时期
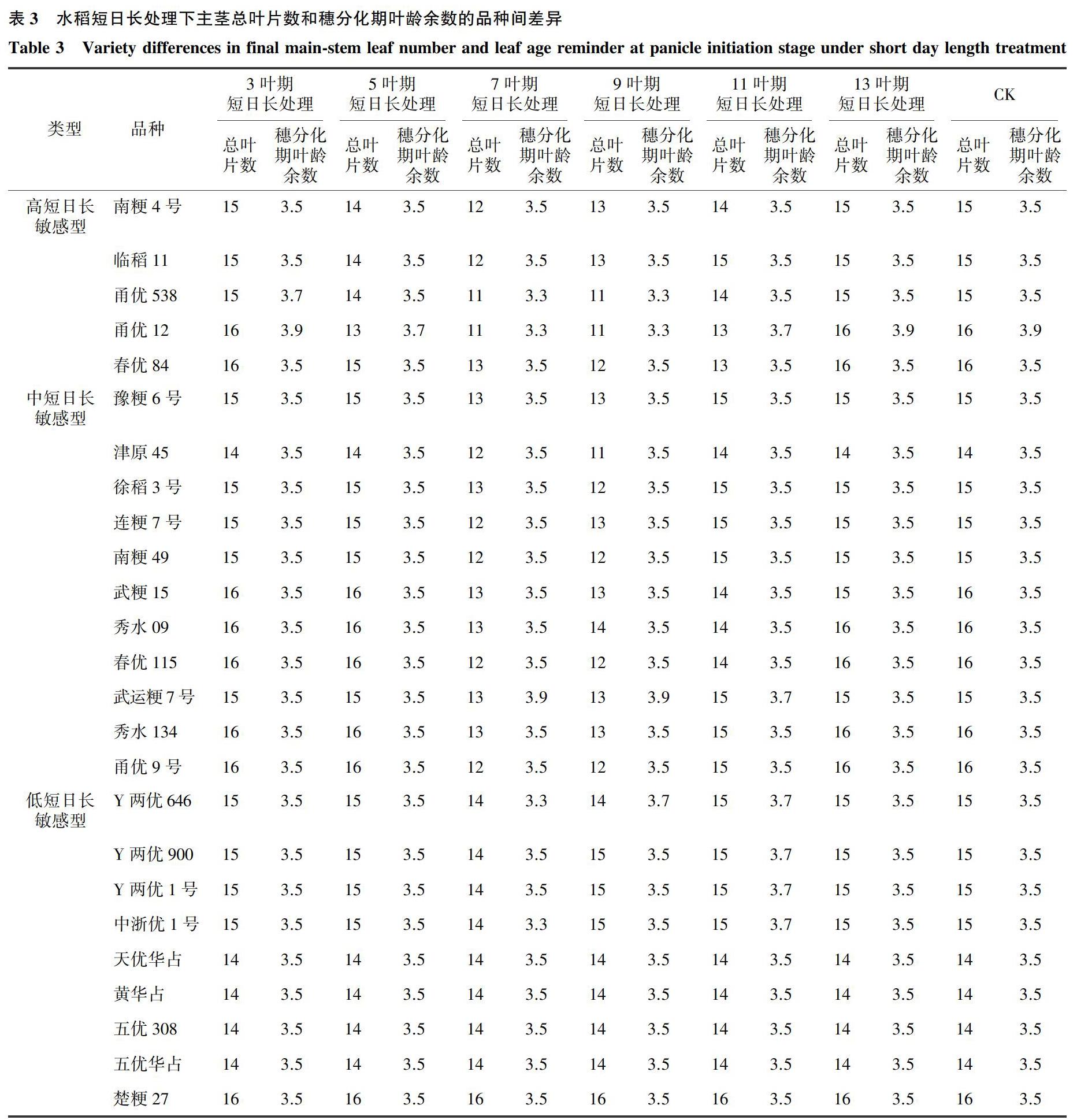
供试水稻品种在正常条件下,主茎总叶片数为14~17叶。供试水稻品种不同叶龄期短日长处理和对照的平均主茎总叶片数见图1。对照的主茎总叶片数最多,除3叶期短日长处理外,其他处理的总叶片数均有不同程度的下降,其中,7~9叶期短日长处理下,总叶片数最少,平均减少叶片数2.0张,且变异系数最大(表2)。水稻不同叶龄期短日长处理下,穗分化期叶龄及总叶片数存在较大变异(表2)。对照和3叶期短日长处理的穗分化期叶龄基本一致,其他叶龄期短日长处理的叶龄较对照不同程度缩短。其中,7~9叶龄期短日长处理,穗分化期叶龄较对照平均提早2.0~2.1叶,但变异系数最大。短日长处理虽然对穗分化期叶龄造成影响,但穗分化期叶龄倒叶数变化不大,平均在倒3.5叶左右,受短日长处理影响不大。以上结果表明,水稻主茎总叶片数对短日长处理的敏感时期在7~9叶期间,穗分化期叶龄倒叶数受短日长影响不大。
2.2水稻主茎总叶片数对短日长处理响应的品种间差异
根据短日长处理下的各水稻品种主茎叶片数变化,利用最长距离法进行聚类分析,结果表明,可将供试品种分为高短日长敏感型,中短日长敏感型和低短日长敏感型3类。3类中,各品种不同叶龄期短日长处理下主茎总叶片数和穗分化期叶龄余数变化见表3。低短日长敏感型品种不同叶龄期短日长处理对主茎总叶片数影响不大。对于中短日长敏感型和高短日长敏感型来说, 3叶期、5叶期、11叶期和13叶期处理主茎总叶片数和对照相比没有显著差异,7叶期短日长处理主茎总叶片数要大于9叶期处理。对于穗分化期叶龄余数来说,不同类型差异不大。
2.3水稻主茎总叶片数对短日长处理响应的品种类型间差异
根据供试品种类型,将所有品种分为籼稻、粳稻和籼粳杂交稻3种类型。3种类型品种主茎总叶片数在不同叶龄时期短日长处理的总叶片数见图2。与对照相比,从5叶期开始,短日长处理不同程度地导致主茎总叶片数减少,粳稻和籼粳杂交稻品种下降幅度最为明显,籼稻品种减少0.1张,其中7叶期和9叶期短日长处理主茎总叶片数减少幅度最大。结果表明,籼粳杂交稻品种的总叶片数对短日长处理最敏感,其次为粳稻,籼稻最弱。
3讨论
水稻主茎总叶片数受日长的影响,短日长处理会减少水稻主茎叶片数[18]。水稻生育期不同阶段对日长的响应不一,水稻需要适宜的短日长处理才能进行穗分化[16],孟亚利等[15]指出水稻从4叶期开始感光。本研究发现,生产中主茎总叶片数为14~17叶的水稻,在7~9叶期对短日长最敏感(图1),短日长处理减少了穗分化期的叶龄,而水稻穗分化整个时期的叶片数并没有受到大的影响,说明水稻生长达到一定叶龄后才开始感光,这和唐锡华等[19]报道相类似。Ritchie等[20]指出,短日长处理会对水稻穗分化期造成影响,但试验发现不同短日长处理水稻穗分化整个时期的叶片数集中在3~4片叶,叶片数的变化和凌启鸿等[4]报道的叶龄模式是一致的,即水稻在主茎倒4叶至倒3叶之间时开始穗分化。研究结果说明,利用叶片数对穗分化期进行判定是准确的。然而,研究发现3叶期短日长处理使水稻抽穗期有所延迟,这可能和短日长处理打乱了植株的碳氮代谢的生理平衡有关[21]。7叶和9叶期短日长处理,高短日长敏感型品种叶片数减少幅度大于中短日长敏感型品种,而低短日长敏感型品种叶片数基本没有变化。水稻品种叶片数的差异由于出叶速率和出叶间隔的不同而不同[22-25],Lee 等[8]指出,短日长处理会降低水稻的出叶速率,这可能是不同短日长敏感型品种叶片数受短日长影响的主要原因。
试验供试品种总叶片数为14~17叶,当然一些品种叶片数少于14叶,它们的短日长敏感时期是否还是7叶至9叶叶龄期值得进一步研究,然而这些品种多是早稻,以籼稻为主,其光周期敏感性较弱[12]。生产中一些晚稻品种叶片数也少于14叶,但这些品种多为单季稻和双季晚稻共用品种,比如试验中的YY538,其总叶片数弹性较大,受日长的影响,作晚稻种植时主茎总叶片数随着播种期的推迟而减少。
在不同类型品种中,籼粳杂交稻对短日长表现出最强的敏感性,籼稻对短日长的敏感性最弱。Yin 等[13]发现籼稻的基本营养生长期要比粳稻长,因此表现出较弱的光周期敏感性。籼粳杂交稻生育期为150~170 d,表现出极大的超亲优势,较强的光周期敏感性能够保证水稻在生产中安全齐穗成熟。魏祥进等[26]指出,等位基因E1在延长高光周期敏感性品种生育期和水稻基本营养生长期中起重要作用,另外Ehd1和RFT1等基因在控制水稻抽穗中也起到了重要作用,并在短日长处理后显著表达[27-28]。在粳稻品种中,楚粳27品种表现为对短日长反应不敏感,这和楚粳27种植的区域有关,楚粳27适应云南高海拔地区种植[29],感温性较强,这也说明,水稻品种的光周期敏感性和水稻亚种的分类并没有明显的相关性。
参考文献:
[1]陈峰,赵庆雷,尹秀波,等. 山东省直播稻的发展现状及对策[J]. 山东农业科学,2018,50(5):156-159.
[2]陈波,周年兵,郭保卫,等. 南方稻区“籼改粳”研究进展[J]. 扬州大学学报(农业与生命科学版), 2017, 38(1) :67-72.
[3]龚金龙,邢志鹏,胡雅杰,等. “籼改粳”的相对优势及生产发展对策[J]. 中国稻米, 2013, 19(5): 1-6.
[4]凌启鸿,苏祖芳,张洪程,等.水稻品种不同生育类型的叶龄模式[J]. 中国农业科学, 1983, 16(1): 9-18.
[5]松岛省三. 稻作的理论与技术[M]. 庞城,译. 北京:农业出版社,1966: 121-133.
[6]BROOKING I R, JAMIESON P D, PORTER J R. The influence of daylength on final leaf number in spring wheat[J]. Field Crops Research, 1995, 41: 155-165.
[7]RIGGI E, SCORDIA D, CUZZOLA C F, et al. Leaf appearance rate and final main stem leaf number as affected by temperature and photoperiod in cereals grown in Mediterranean environment[J]. Italian Journal of Agronomy, 2017, 12(3):183-191.
[8]LEE C, LEE B, SHIN J, et al. Heading date and final leaf number as affected by sowing date and prediction of heading date based on leaf appearance model in rice[J]. Korean Journal of Crop Science, 2001, 46: 195-201.
[9]馮阳春,魏广彬,李刚华,等. 水稻主茎出叶动态模拟研究[J]. 中国农业科学,2009,42(4), 1172-1180.
[10]叶宏宝,孟亚利,汤亮,等.水稻叶龄与叶面积指数动态的模拟研究[J]. 中国水稻科学, 2008, 22(6): 625-630.
[11]朱德峰,章秀福,许立,等. 水稻主茎叶片出生与温度关系[J]. 生态学杂志, 1998, 17(5): 71-73.
[12]唐锡华,倪彭寿,童本仙,等. 在控制条件下对不同稻种日长和温度反应发育特性的研究[J]. 植物生理学报, 1978 (4): 153-168.
[13]YIN X Y, KROPFF M J, YNALVEZ M A. Photoperiodically sensitive and insensitive phases of pre-flowering development in rice[J]. Crop Science, 1997, 37(1): 182-190.
[14]YABUTA S, HSKOYAMA S, INAFUKU S, et al. The photoperiodic sensitivity estimation at three growth phases of rice fractionated by turning point of leaf emergence and panicle initiation[J]. Japanese Journal of Crop Science, 2015, 84(1): 64-68.
[15]孟亚利,曹卫星,周治国,等. 基于生长过程的水稻阶段发育与物候期模拟模型[J]. 中国农业科学, 2003, 36(11): 1362-1367.
[16]NOGUCHI Y, NAKAJIMA T, YAMAGUCHI T. Studies on the control of flower bud formation by temperature and daylength in rice plants: IX. Aging effect in relation to photoperiodic induction[J]. Japanese Journal of Breeding, 1971, 21(2): 81-86.
[17]HAUN J R. Visual quantification of wheat development[J]. Agronomy Journal, 1973, 65: 116-119.
[18]YIN X Y, KROPFF M J. Use of the Beta function to quantify effects of photoperiod on flowering and leaf number in rice[J]. Agricultural & Forest Meteorology, 1996, 81: 217-228.
[19]唐锡华,李文安. 水稻光照发育阶段的研究Ⅱ.几种内外条件对水稻光照阶段发育的影响[J]. 作物学报, 1964, 3(3): 283-296.
[20]RITCHIE J T. Systems approaches for agricultural development[M]. Netherlands: Springer Nature,1993:77-93.
[21]韩天富,王金陵. 大豆开花后光周期反应的研究[J].植物学报, 1995,37(11): 863-869.
[22]ERMINI M L, MARIANI S, SCARANO S, et al. Rice crop duration and leaf appearance rate in a variable thermal environment. II. Comparison of genotypes[J]. Field Crops Research, 1998, 58: 129-140.
[23]ITOH J I, HASEGAWA A, KITANO H, et al. A recessive heterochronic mutation, plastochronl, shortens the plastochron and elongates the vegetative phase in rice[J]. The Plant Cell,1998, 10: 1511-1521.
[24]SALAM M A, MACKILL D J. Effects of a photoperiod sensitivity gene on morphological characters in rice[J]. Field Crops Research, 1993,33:261-268.
[25]钟代彬,罗利军,梅捍卫,等. 水稻主茎总叶片数及其相关性状的QTL分析[J]. 中国水稻科学, 2001, 15(1): 7-12.
[26]魏祥进,徐俊锋,江玲,等. 我国水稻主栽品种抽穗期多样性的遗传分析[J]. 作物学报, 2012, 38(1):10-22.
[27]CHO L H, YOON J, PASRIGA R, et al. Homodimerization of Ehd1 is required to induce flowering in rice[J]. Plant Physiology, 2016,170: 2159-2171.
[28]KOMIYA R, IKEGAMI A, TAMAKI S, et al. Hd3a and RFT1 are essential for flowering in rice[J]. Development, 2008, 135: 767-774.
[29]李开斌,张天春,阮文忠,等.超级粳稻楚粳27号的选育及应用[J]. 中国稻米, 2012, 18(5): 71-72.
(責任编辑:陈海霞)
