Vegetation structure, dominance patterns and height growth in an Afromontane forest, Southern Africa
2020-05-22SylvanusMensahAnthonyEgeruAchilleEphremAssogbadjoRomainGleleKaka
Sylvanus Mensah · Anthony Egeru · Achille Ephrem Assogbadjo ·Romain Gle`le` Kakaı¨
Abstract Information on forest structure is fundamentally important to track successional vegetation dynamics for efficient forest management. This study reports on vegetation characteristics, dominance patterns and species height growth in a northern mistbelt forest type in South Africa. Common alpha-diversity indices (species richness and Shannon-Weiner diversity), structural vegetation parameters (tree density and basal area), and species importance value index were used. Size class distribution and height-diameter allometry were further examined for the overall stand and most important species. Stem densities (472.0 ± 43.5 and 605.3 ± 28.1 trees ha-1 for ≥5 cm to <10 cm and ≥10 cm dbh (diameter at breast height) classes, respectively) and basal area values(1.99 ± 0.19 and 48.07 ± 3.46 m2 ha-1, respectively) are comparable to other Afromontane forests in East Africa.The overall stand showed an inverted-J shaped distribution pattern which is a typical feature of stand size class distribution in most natural forests. Most ecologically important species also exhibited an inverted-J shaped distribution pattern, suggesting good regeneration and recruitment potential. There were significant differences in species on height, reflecting species-specific height growth patterns, possibly a result of intrinsic growth potential and competitive interactions. The present study suggests that conservation and management policies, including protection of surrounding land uses against fire, contribute to maintaining a successful recovery of these forests. However, it should be noted that these forests may be experiencing relatively slow dynamic flux as a result of the overmature state of some trees with several years under relatively strict protection.
Keywords Diversity · Population structure · Species composition · Size class distribution
Introduction
Over the last century, tropical forests suffered severely from natural disturbances (fires, winds, floods) and timber harvesting. Logging and forest clearing for subsistence agriculture increase loss of biodiversity, which affects ecosystem integrity, functions and services (Foley et al.2007). South Africa has not been immune to such impacts(King 1941; Cooper 1985), as the country’s forests have been exposed to fire and unregulated harvesting. Nevertheless, by 1939, illegal logging was prohibited, and this,added to the efforts to control fire, has contributed to the restoration of natural forest vegetation in many degraded areas (Geldenhuys 2002).
Afromontane mistbelt forests are one of the few natural forest ecosystems in South Africa which, due to the modification of the fire regime, have developed in areas that were not previously covered in forests (Geldenhuys 2000;Geldenhuys and Venter 2002). These forests persisted in a fire-prone environment and then expanded into suitable areas with fire protection for timber plantations and intensive agriculture. They consist of many small, fragmented and widely distributed patches. Two mistbelt forests form part of the eight natural forest groups in South Africa: the northern mistbelt forests and the southern mistbelt forests(Mucina and Rutherford 2006). One of the most striking characteristics in the northern mistbelt forests is the tall moist evergreen vegetation at altitudes up to 1800 m.These forests serve as habitats for insect pollinators(Mensah et al. 2017b), but also play important socio-economic and ecological roles that support human well-being.Among other benefits, they provide forage resources to domestic animals and insect pollinators (contributing to crop pollination in marginal agricultural areas), fuel wood,food, medicinal resources, help to control biological invasions, and alleviate the effects of climate change by storing atmospheric carbon (Geldenhuys 2002; Rasethe et al. 2013;Mensah et al. 2016a, 2017a).
Information on current floristic composition and species dominance are important to track successional dynamics of the forest vegetation and its response to environmental variations and human interventions. Most previous research in the northern mistbelt forests assessed the potential for regeneration and traditional plantation forestry of native species (Geldenhuys and von dem Bussche 1997; Geldenhuys 1997), growth and mortality of indigenous species(Geldenhuys 2000), and biodiversity and floristic composition for management options (Geldenhuys and Venter 2002).Most of these studies were conducted more than a decade ago and although diversity and vegetation patterns are known to be responsive to environmental conditions, species distribution patterns, recovery and growth rates can also shift on a temporal scale in response to the nature and intensity of surrounding land uses, and to management practices (Kennard 2002; Sampaio et al. 2007). Information on current forest structure (horizontal and vertical) are crucial to characterize the vegetation, and describe and interpret their dynamics. This information is also relevant to appreciate the effectiveness of management interventions in surrounding land uses as well as conservation decisions.
In this study, recently collected inventory data from the Woodbush-De Hoek natural forest in Limpopo province,South Africa was used to explore patterns of species diversity, vegetation structure and dominance. The specific objective was to assess (i) overall vegetation patterns (including dominance), focusing on taxonomic diversity metrics (species richness and Shannon-Weiner diversity index), and (ii) common vegetation structural parameters such as regeneration density, tree density, basal area,Importance Value Index, and size class distribution.
Materials and methods
Study area and data
The Woodbush-De Hoek natural forest is situated at 23°50′S and 29°59′E and is part of the northern mistbelt forest group. The vegetation is dominated by tall evergreen species at altitudes up to 1800 m (Geldenhuys 2002). The area is surrounded by pine plantations managed by the Komatiland Forests Company in Limpopo. The mean annual rainfall is 1200 mm, with the highest peak during summer. Additional information about the study area and Woodbush-De Hoek natural forest are available in our previous studies (Mensah et al. 2018a, b).
Floristic and structural data were collected by means of forest inventories. Thirty circular plots of 500 m2each were randomlylaid out and thirtyadditional circular subplots(250 m2) were laid out in each 500 m2plot. Inside the subplots, diameter at breast height (dbh) was measured for all stems ≥5 cm to <10 cm while in the 500 m2plots, the same measurements were taken for all trees with dbh ≥10 cm. Both dbh sizes were recorded to gain more insight into regeneration and adult trees. Species names were recorded using taxonomic keys of the revised and updated version of the book, Trees of Southern Africa (Coates-Palgrave 2003).
Data analysis
Taxonomic diversity and floristic composition were analysed for all woody plants having dbh ≥5 cm. Taxonomic diversity, species richness and Shannon-Weiner diversity index were computed following Magurran (1988). Structural variables such as stem density (N, trees ha-1) and basal area (G, m2ha-1) were computed and combined to provide relative species frequencies to determine the importance value index (IVI; Curtis and McIntosh 1951) as an indicator of a species relative ecological importance.Tree density was calculated as the average number of trees per hectare, and basal area as the sum of the cross-sectional area 1.3 m above the ground using the following equation:

where G is basal area; diis diameter in cm of the i-th tree in the plot and s the unit area of the plot. IVI was calculated by summing up the species relative frequency, relative density and relative basal area as:
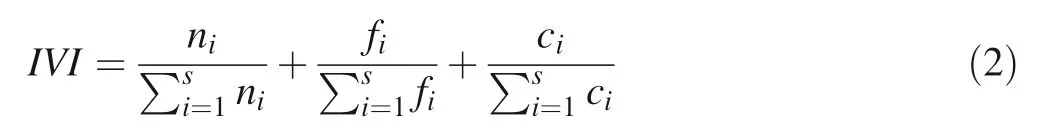
where IVI is the importance value index; ni, fiand ciare respectively the density, frequency and basal area of the ith species. Size class distribution and height growth models are important in understanding the dynamics of forest stands. Stem diameter structures were established for the overall stand and the ten most important species as revealed by IVI. To better depict the size class distribution patterns, we used the method of Condit et al. (1998). For each species, we performed a least square linear regression analysis using the number of individuals as dependent variable and the midpoint of each size class as independent variable. Each size class had 10 cm width, starting from 5 cm dbh. Because some size classes had no recording, the number of individual species in each class was transformed by ln (1 + number of individuals), as suggested by Obiri et al. (2002). The slope from each species-specific size class regression was used as an indicator of the level of recruitment and population structure (Lykke 1998). Negative slope values are indicative of a reverse-J SCD (size class distribution) curve and of good recruitment, with more individuals in smaller size classes and fewer in larger classes (Obiri et al. 2002; Martins and Shackleton 2017).Positive slope values indicate populations with a unimodal SCD curve and limited recruitment, with more individuals in larger size classes and fewer in smaller size classes.
For height growth, seven out of the ten most important species were considered due to the limitation of height data. The allometric relationship between tree diameter and total height for these species were examined using the linearized form of the power function (Mensah et al.2016a, 2017c). We tested whether height-diameter relationship varied among species by adding the species as a categorical factor and performing an analysis of covariance. Individual and interaction term effects were computed from the fitted model:

where, ln (α) is the intercept, β the proportional height growth rate induced by tree diameter, and γspeciesthe proportional growth rate due to the species; ln (dbh) is the natural logarithm of tree diameter and ε′the additive error.
Results
Diversity, structures and dominance patterns
Fifty woody plant species belonging to 46 genera and 33 families were recorded. The overall Shannon-Wiener diversity was 2.84. Species richness was slightly lower for ≥5 cm to <10 cm dbh class (35 species), compared to the ≥10 cm dbh class (45 species). However, further analyses showed that the number of species per plot was significantly higher for smaller trees (<30 cm dbh) and lowest for larger trees (>60 cm) (F = 115.19; P <0.001;Fig. 1). The most diversified families were the Rutaceae and Rubiaceae, with five and four species, respectively.
In terms of relative basal area, the most dominant families were the Myrtaceae (12.7%) and Monimiaceae(8.3%) (Table 1). These two families also had the highest number of individuals, although both were represented by only three species (Xymalos monospora (Harv.) Baill,Syzygium gerrardii (Harv. ex Hook. f.) Burtt Davy and Eugenia natalitia Sond.). Stem densities of the ≥5 to <10 cm and ≥10 cm dbh classes were 472.0 ± 43.5 and 605.3 ± 28.1 trees ha-1, respectively. Xymalos monospora, Syzygium gerrardii, Combretum kraussii Hochst.,Cussonia sphaerocephala Strey, and Cassipourea malosana (Baker) Alston were the most abundant species in the ≥10 cm dbh class, with 139.3, 119.3, 43.3, 30.7 and 24.7 stems ha-1, respectively. For the ≥5 to <10 cm dbh class, Cassipourea malosana (68.0 stems ha-1), Kraussia floribunda Harv. (56.0 stems ha-1), Xymalos monospora(50.7 stems ha-1), Syzygium gerrardii (44.0 stems ha-1),Ochna arborea Burch. ex DC. (36.0 stems ha-1) and Rinorea angustifolia (Thouars) Baill. (34.7 stems ha-1) had the highest densities, accounting for 61.3% of the density in this class.
The basal areas for the ≥5 to <10 cm and ≥10 cm dbh classes were 1.99 ± 0.19 and 48.07 ± 3.46 m2ha-1,respectively. Out of the 50 species, 42 had basal areas less than 1.0 m2ha-1, contributing less than 20.0% of the stand basal area; five species had basal areas between 1.0 and 3.0 m2ha-1(20.0% of the stand basal area). The greatest contribution to stand basal area (>25%) was made only the forest waterberry (Syzygium gerrardii (Fig. 2). The ten most important species based on the Importance Value Index were, in order of ecological importance: X. monospora, S. gerrardii, C. sphaerocephala, C. kraussii, C.malosana, Trichilia dregeana Sond., Nuxia congesta R. Br.ex Fresen., Kraussia floribunda, Croton sylvaticus Hochst.,and O. arborea (Table 2).

Fig. 1 Variation in plot species richness among larger, medium-sized and smaller trees
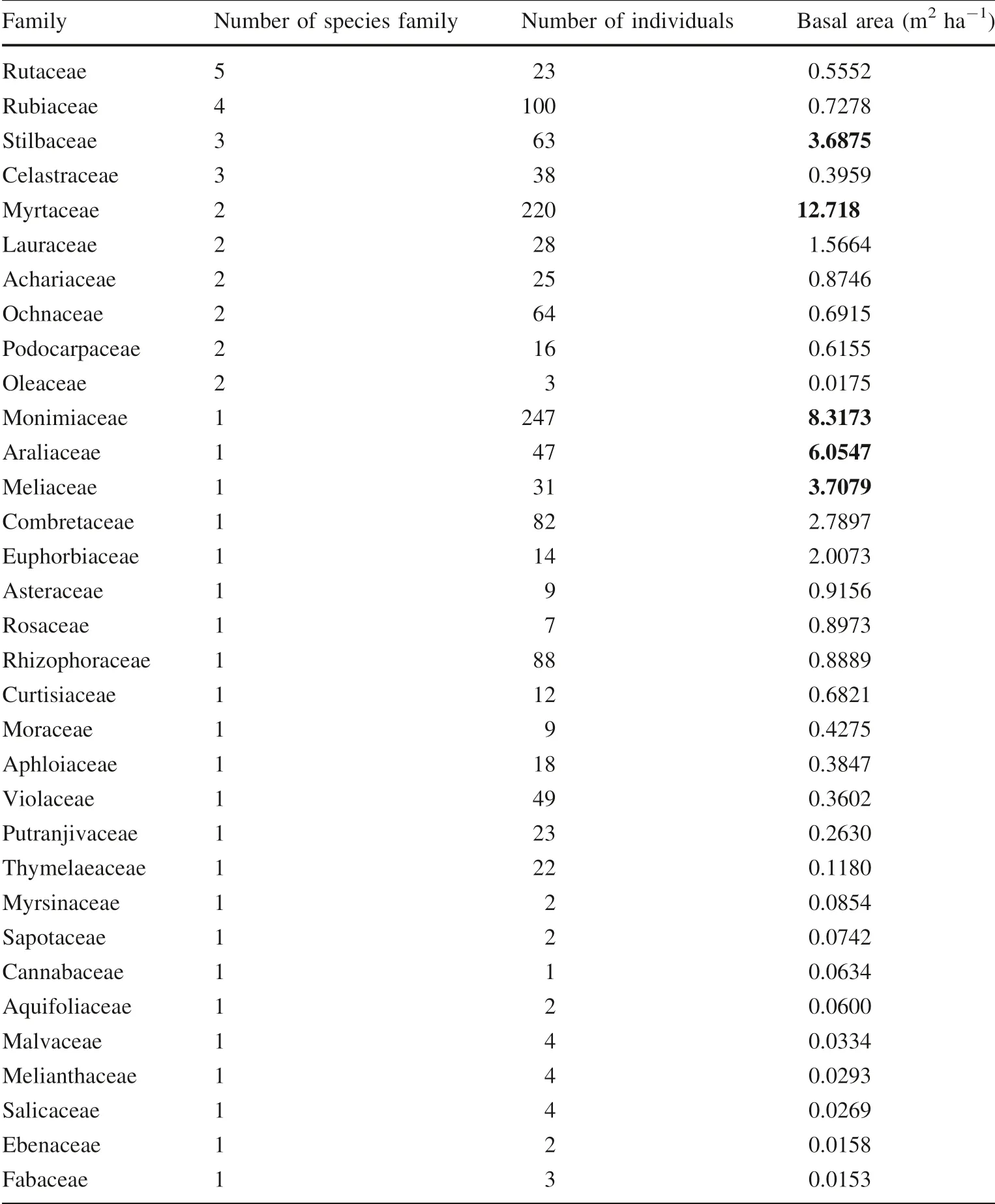
Table 1 Number of species,individuals and basal area per taxonomic family
Size class distribution
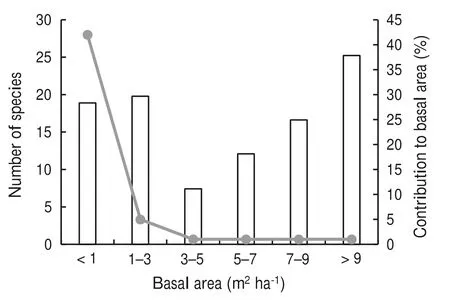
Fig. 2 Variation in number of species (solid line) according to their contribution to the stand basal area

Table 2 Species names and importance value index (IVI)
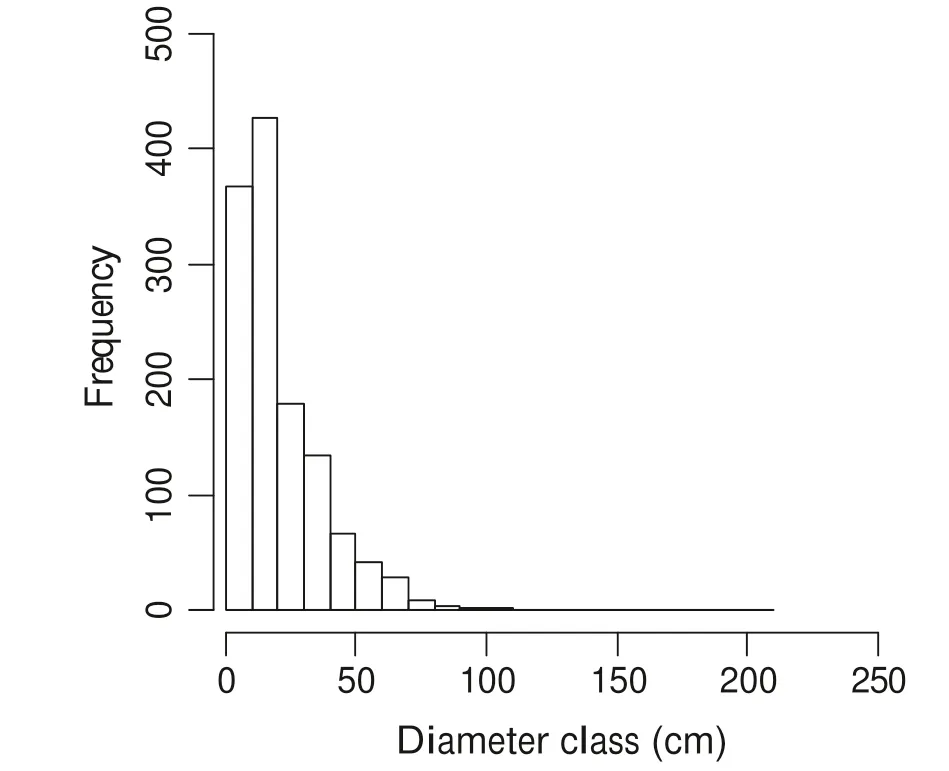
Fig. 3 Size class distribution for the overall stand
The overall size class distribution showed an inverted-J shaped curve with several gaps in diameter classes beyond 100 cm dbh (Fig. 3). Overall, most trees were in the lower diameter classes (<20 cm), and shows a drastic decline in density in the higher classes. Diameter class structures were further established for the ten most important species(Fig. 4). All species exhibited a tendency to the inverted J-shape distribution, indicating higher frequency in the lowest diameter classes and a gradual decrease from lower to higher size classes. The J-shape distribution was more pronounced for X. monospora, S. gerrardii, C. kraussii, C.malosana, O. arborea and K. floribunda. C. sphaerocephala, T. dregeana, N. congesta and C. sylvaticus showed a size class distribution with several gaps in intermediate size classes. From the results of regression analyses of SCD curves, these species had negative and steeper slopes (from - 0.09 to - 0.04; Table 3), as compared to N. congesta (slope = - 0.03), T. dregeana(slope = - 0.01), C. sphaerocephala (slope = - 0.01) and C. sylvaticus (slope = - 0.009). Size class distribution slopes for both O. arborea and K. floribunda were not significant due to the low number of size classes (Table 3).These slope values suggest that species such as X.monospora, S. gerrardii, C. kraussii, C. malosana had a higher number of individuals in smaller size classes.Meanwhile, species such as C. sphaerocephala, S. gerrardii and C. kraussii were still present in the higher diameter classes (>80 cm).

Fig. 4 Size class distribution for the ten most important species

Table 3 Slope of species-specific size class distribution
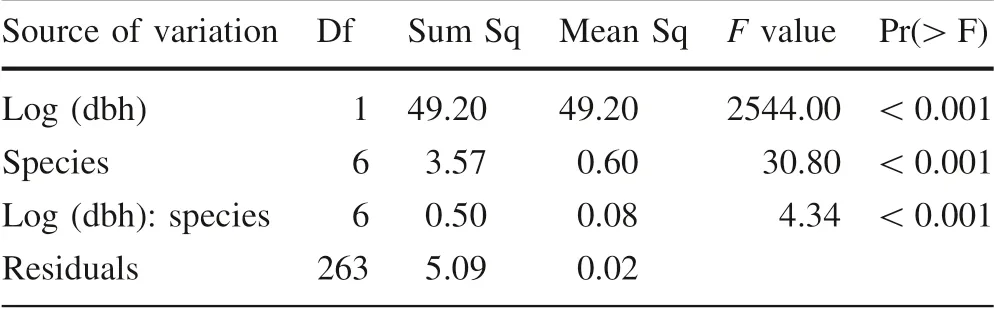
Table 4 Results of the analysis of covariance on the effects of species and diameter on height
Dominant species height-diameter allometry
The results of the analysis of covariance show significant effects of diameter and species on height growth (Table 4).There was also significant interaction between diameter and species (F statistic = 4.34; p <0.001; Table 4), indicating that height growth varied among dominant species.As expected, height increased with increasing diameter for the seven dominant species (Table 5). For a given dbh, C.malosana and C. sylvaticus had slopes that were respectively 0.27 ± 0.10 and 0.13 ± 0.06 significantly higher than the ones for K. floribunda, S. gerrardii, T. dregeana,X. monospora and C. kraussii, which was used here as a reference. These results indicate that, for the same dbh, C.malosana and C. sylvaticus had significantly higher average heights at tree level, as compared to K. floribunda, C.kraussii, S. gerrardii, X. monospora and T. dregeana(Fig. 5).
Discussion
When compared with Afromontane forests in Ethiopia(Kebede et al. 2013; Tadele et al. 2014), our results show,despite the limited spatial coverage, that the northern mistbelt forest harbours considerable floristic diversity in terms of both tree species and taxonomic families. More species were expected to be found in the regeneration class,as general floristic patterns in multispecies natural forests.Out of the 50 species enumerated, 35 were in the ≥5 to <10 cm dbh class and 45 in ≥10 cm dbh class. The slightly lower species richness in the 5-0 cm class could be due to the 5-cm threshold considered, which excludes additional species <5 cm. Our results further reveal thatspecies richness decreased with increasing size class(Fig. 1). Other studies have also demonstrated that species richness declines from lower to upper diameter classes(Lulekal et al. 2008; Kebede et al. 2013). Our findings are also in agreement with Guilherme et al. (2004) who reported higher floristic richness and diversity in lower/intermediate layers than in upper layers of tropical forests.The diversity patterns observed in regards to diameter classes may be the results of species interactions (especially competition for light), which also determines the maximum height.
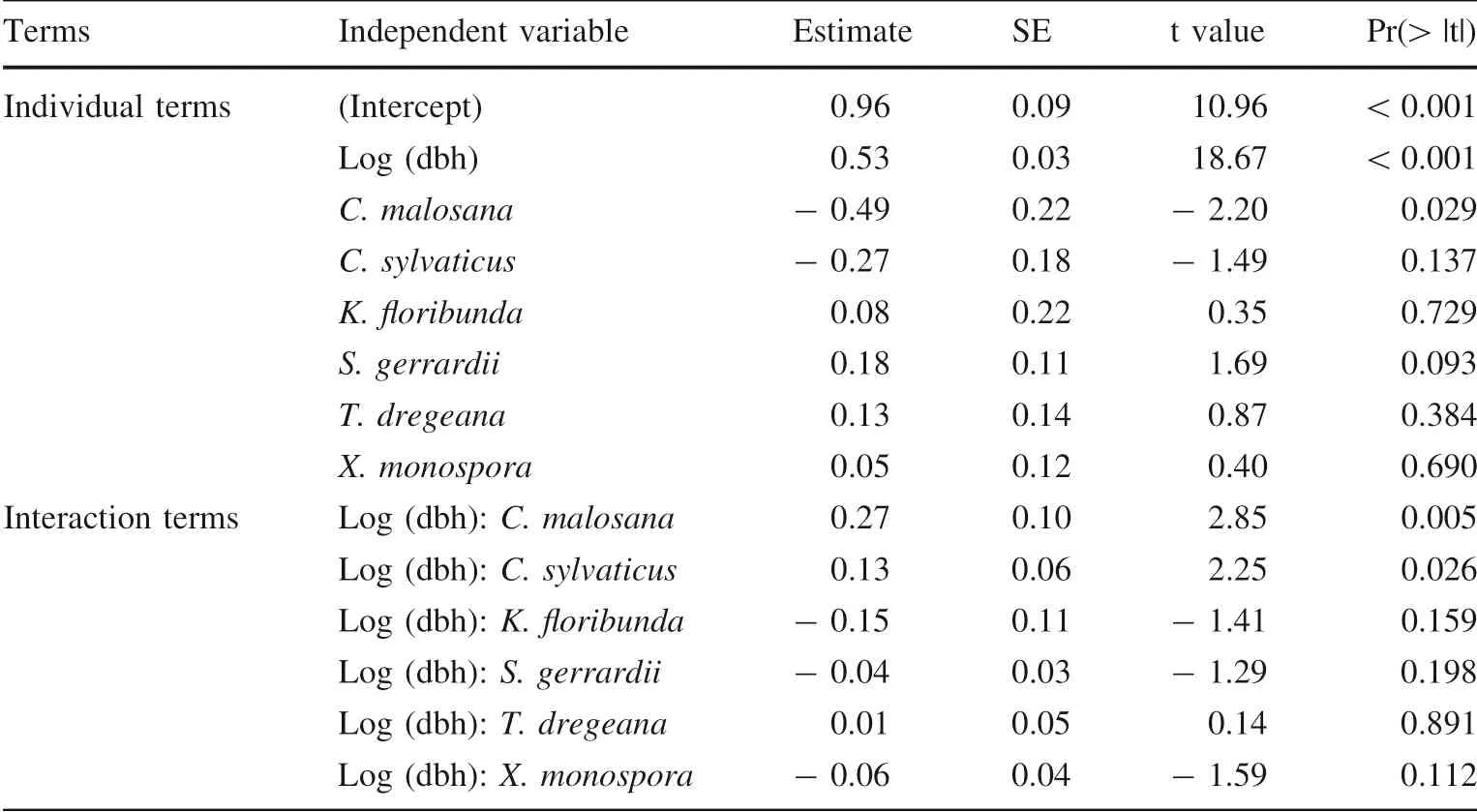
Table 5 Results of the linear models of individual and interaction effects of species and diameter on height

Fig. 5 Height-diameter allometries for important species. Scatter plots are based on 277 trees from seven dominant species
Tree density and basal area were respectively,472.0 ± 43.5 stems/ha and 1.99 ± 0.19 m2ha-1for the≥5 to <10 cm dbh class, and 605.3 ± 28.1 stems/ha and 48.07 ± 3.46 m2/ha for the ≥10 cm class, suggesting that these forests have great timber potential, comparable to other Afromontane forests in East Africa (Lulekal et al.2008; Fisaha et al. 2013; Kebede et al. 2013). For example,with 75 (20 m × 20 m) quadrats recording diameters and heights greater than 2 cm and 2 m, respectively, Kebede et al. (2013) reported lower values of tree density (379.3 stems/ha) and basal area (31.4 m2ha-1) in a remnant moist Afromontane forest of Wondo Genet in south central Ethiopia. Yirdaw et al. (2015) reported similar density values (537.0-622.0 stems ha-1) and relatively lower basal area (29.4-36.2 m2/ha) in Afromontane cloud forests in the Bale Mountains of southeast Ethiopia. Knowing that these structural features are key indicators of forest biomass potential, our results partly confirm the remarkable biomass and carbon potential of these mistbelt forests (Mensah et al.2016a). Over past decades, wild fires and forest fragmentation took significant toll on these indigenous forests, now confined to small, scattered patches. Given the historical disturbances, it is important to note that conservation policies and recent management interventions, including the protection of surrounding land uses against fire, have contributed to successful development of these forests. X.monospora and S. gerrardii were the most abundant and dominant species in the inventoried forest area. These two species contributed more than 40% of the stand basal area.Past studies in the Limpopo mistbelt forests also showed similar dominance patterns (Geldenhuys 2002; Geldenhuys and Venter 2002).
The overall size class distribution was an inverted J-shape with a sharp decline in tree density towards the higher classes, a typical feature of diameter class distributions in most Southern Africa’s forests (Geldenhuys 1993, 1996; Mujuru and Kundhlande 2007; Kebede et al.2013). Few individual trees were over 80 cm, as also shown in previous reports by Geldenhuys (2000) and Geldenhuys and Venter (2002). The overall distribution is characteristic of multi-species forest stands with sufficient regeneration. An inverted J-shape without gaps is also indicative of a species successful reproduction and succession in diameter classes. Inverted J-shape trends were observed for X. monospora, S. gerrardii, C. kraussii, C.malosana, O. arborea, and K. floribunda, indicating that their regeneration pool would be sufficient for maintaining their populations. However, given the 5-cm diameter threshold considered in this study, more in-depth analyses on stability across diameter classes for these species would provide insights into their ecological status and dynamic patterns. Species such as C. sphaerocephala, T. dregeana,N. congesta and C. sylvaticus exhibited several gaps in lower and intermediate size classes, which may be partially due to natural events such as tree fall. For instance, the gaps in C. sphaerocephala diameter class distribution is possibly the result of frequent wind throw on steep slopes and/or trees dying. Gaps in lower and intermediate size classes may also be attributed to irregular and sporadic seedling establishment as a result of competition for nutrients and light (Geldenhuys 1993; Syampungani et al.2015). When examining the relationship of species to height, it was interesting to see that species with relatively lower wood density such as C. sylvaticus (Mensah et al.2016b), tend to have taller individuals. This is partially attributable to the fact that species with lower wood density grow faster than those with higher wood density (Mensah et al. 2016a, b). The variation in height growth between dominant species is also indicative of species-specific performance resulting from differential resource acquisition, competition and biomass production.
Limitations and conclusion
We assessed the patterns of species diversity, vegetation structures and dominance in a northern mistbelt forest type in South Africa. Vegetation observational studies often benefit from larger sample and plot size. The sample size used in this study was slightly lower than what Yirdaw et al. (2015) (n = 36; 400 m2) used in studying floristic diversity and structure in Afromontane cloud forests in Ethiopia. Thus, our sampling design might have not captured the overall variation in these forests composition.Despite the limited spatial coverage of our data, the results in terms of flora and structure were comparable to previous reports in other Afromontane forests. The overall stand showed an inverted-J shaped diameter distribution pattern,a typical feature of size class distribution in natural forests;and similar diameter structures were observed for most dominant species. We also found significant effects of species on tree height, suggesting differential height growth among dominant species, probably as result of different wood density and competitive interactions.
Overall, this study suggests that the long period of relatively strict conservation and the management policies,including protection of surrounding agricultural and forestry land uses against fire, have contributed to maintain a successful recovery of the forests. Considering adverse climate and land use change effects, our results indicate that diversity and structure in the study area will generally benefit current from legal protection. However, these forests might be experiencing a relatively slow dynamic flux as a result of minimal anthropogenic disturbance and the over-mature state of larger trees. Institutional and managerial policies permitting few extractive human uses would contribute to preserve a healthy ecosystem, which is in line with the intermediate disturbance hypothesis and the general idea that anthropogenic disturbance cannot be abandoned as a regulatory force in species structuring(Huston 2014).
AcknowledgementsThe data used in this study were collected during the first author’s doctoral research field work, co-financed by the African Forestry Forum and the National Research Foundation of South Africa through the ‘‘Catchman Letaba’’ project. The authors are grateful to the anonymous reviewers for the comments on the first version of this paper.
杂志排行

Journal of Forestry Research的其它文章
- Past, present and future of industrial plantation forestry and implication on future timber harvesting technology
- Effects of climate changes on distribution of Eremanthus erythropappus and E. incanus (Asteraceae) in Brazil
- Effects of climate and forest age on the ecosystem carbon exchange of afforestation
- Effect of gap size and forest type on mineral nitrogen forms under different soil properties
- Effect of forest thinning on hydrologic nitrate exports from a Nsaturated plantation
- Floristic analysis and dominance pattern of sal (Shorea robusta)forests in Ranchi, Jharkhand, eastern India