温度和水流对八斑唇腕水母生长发育的影响
2020-05-13杨翠华王文章王海铭
杨翠华, 王文章, 王海铭
温度和水流对八斑唇腕水母生长发育的影响
杨翠华, 王文章, 王海铭
(青岛海洋科技馆, 山东 青岛 266003)
在不同温度和水流条件下, 作者分别对八斑唇腕水母()伞径、伞高、出芽率和存活时间等生长发育情况展开研究, 以期为其人工饲养和科普展示奠定基础。近海9.5℃~14℃采集的八斑唇腕水母暂养1 d后, 置于不同水流(静止的1 L烧杯和气泡带动水流的13 L水浴缸)和不同温度(10℃、15℃、20℃和22℃)交叉条件下人工饲养。结果显示, 77 cm/s的水流条件下, 10℃组水母存活时间最长。伞径、伞高与饲养时间呈负相关。温度越高, 水母存活时间越短。温度能促进生殖腺快速成熟, 但是10℃水母能持续性出芽。水流能保持水母活力, 延长存活时间。研究表明, 八斑唇腕水母可以通过水螅体横裂生殖和性腺出芽生殖两种方式繁育水母体。因为其个体较小, 科技馆可采用低温水浴缸进行培养和长期展示, 并对其特殊的出芽生殖方式进行科普讲解。
八斑唇腕水母(); 温度; 水流; 出芽; 存活时间
八斑唇腕水母(), 又称八斑芮氏水母, 属水螅虫总纲(Hydrozoa)、水螅水母纲(Hydroidomedusa)、丝螅水母目(Filifera)、唇腕水母科(Rathkeidae)、唇腕水母属()[1]。自然界中, 水母体伞高3 mm~4 mm, 高略大于宽; 伞呈梨形, 有实心顶突; 胃短, 长方形, 具有锥形的胃柄; 生殖腺围绕在胃柄基部[2]。与大多数水螅水母由水螅体产生水母体不同, 八斑唇腕水母不仅可以通过固着的水螅体产生水母体[3](图1), 而且可以通过围绕在胃壁上的生殖腺, 生出水母芽[4](图2), 脱落后发育为可以自由游动的水母体。Bouillon等[5]研究了八斑唇腕水母芽的形成过程, 结果显示, 出芽生殖的水母体仅仅由母体的外胚层产生, 而水螅体产生的水母体来源于水螅体的内胚层和外胚层。两种方式产生的水母体伞高和伞径的比值不同[6]。
八斑唇腕水母与大多数水母一样, 存在世代交替[7]。1950年, Berrill[4]观察到八斑唇腕水母无性出芽生殖。1958年, Werner[3]在室内通过改变饲养温度, 第一次获得了八斑唇腕水母的水螅体。本试验通过在封闭循环缸内饲养发育成熟的水母体, 第一次观察到有性繁育阶段游离的生殖细胞、受精卵和发育过程, 根据实验室观察绘制了生活史示意图(图3)。
由于具有特殊的繁殖方式和优美的外形, 随着对八斑唇腕水母实验室观察的深入和饲养经验的积累, 水族馆对其水母体长期科普展示的需求越来越高。目前研究主要集中在基础分类描述方面[8-9], 关于水温和水流对八斑唇腕水母饲养和繁育的影响未见报道。本文研究了水温和水流对其形态、出芽率和存活时间的影响, 旨在丰富该水母的生物学资料, 以期为采集水螅水母的人工驯化和室内繁育提供必要的参考资料。
1 材料与方法
1.1 试验材料
本试验用水母2016年3月采集于鲁迅公园前海码头, 采集海域表层水温9.5℃~14℃, 用200目筛绢捞起。实验在青岛海洋科技馆实验室内进行, 采集的成熟水母体放在15℃生化培养箱的1 L烧杯内静水暂养。试验用水经过黑暗沉淀、砂滤、臭氧、活性炭预处理, 盐度32~33, pH7.5~8.2。1 d后, 挑选平均伞径3.65 mm、伞高4.08 mm、大小一致的成体水母随机分组饲养。每日10:00投喂孵化24 h的卤虫无节幼体(), 投喂4 h后完全换水。

图1 正在横裂的水螅体

图2 正在出芽的八斑唇腕水母体

图3 八班唇腕水母生活史示意图
1.2 方法
1.2.1 实验设计
水流静止组试验在生化培养箱的1 L烧杯内进行, 水流组在充气带动水流的13 L圆形水浴缸内进行, 采用漂流测速法测定水流速率[10], 重复测定6次, 计算平均值为77 cm/s。试验设10℃、15℃、20℃、22℃共4组, 温差±1℃, 每组放入30个成体水母, 每天完全换水后, 将分裂的水母幼体取出, 最初分组的水母成体继续试验。每3 d用显微镜测量10只水母的伞径和伞高, 记录水母出芽情况(用脱离母体的小水母表示)和存活数。
1.2.2 测定方法
将水母放在载玻片水滴中1 min, 使其充分伸展, 在10×1倍奥林巴斯SZ61体视显微镜下使用cellsens standard1.6软件测量伞径和伞高。
1.3 数据统计与处理
数据统计结果表示为平均值±标准差(Mean±SD), 对数据进行正态分布检验(Kolmogorov-Smirnov test)和方差齐性检验(Levene test)。使用Excel2010统计平均伞径、伞高, 及其标准差, 并对伞径、伞高与饲养时间进行相关性分析。使用SPSS19.0统计软件进行One-way ANOVA分析, Duncan法进行多重比较, 显著性水平为0.05。
2 结果与分析
2.1 温度和水流对伞径、伞高的影响
不同条件下水母伞径、伞高分别见表1和表2。实验用八斑唇腕水母伞径平均3.65 mm,伞高4.08 mm, 静止组伞径和伞高均与饲养时间负相关。10℃、15℃、20℃和22℃组水母在人工饲养后30、24、18、12 d后, 伞径分别为2.68、3.00、2.58和2.60 mm, 第二天均全部死亡。水流组水母伞径变化趋势与静止组类似, 但是能更长时间的保持自然状态。

表1 不同条件下水母伞径
注: 表中的“—”表示水母全部死亡(表2、表3同)
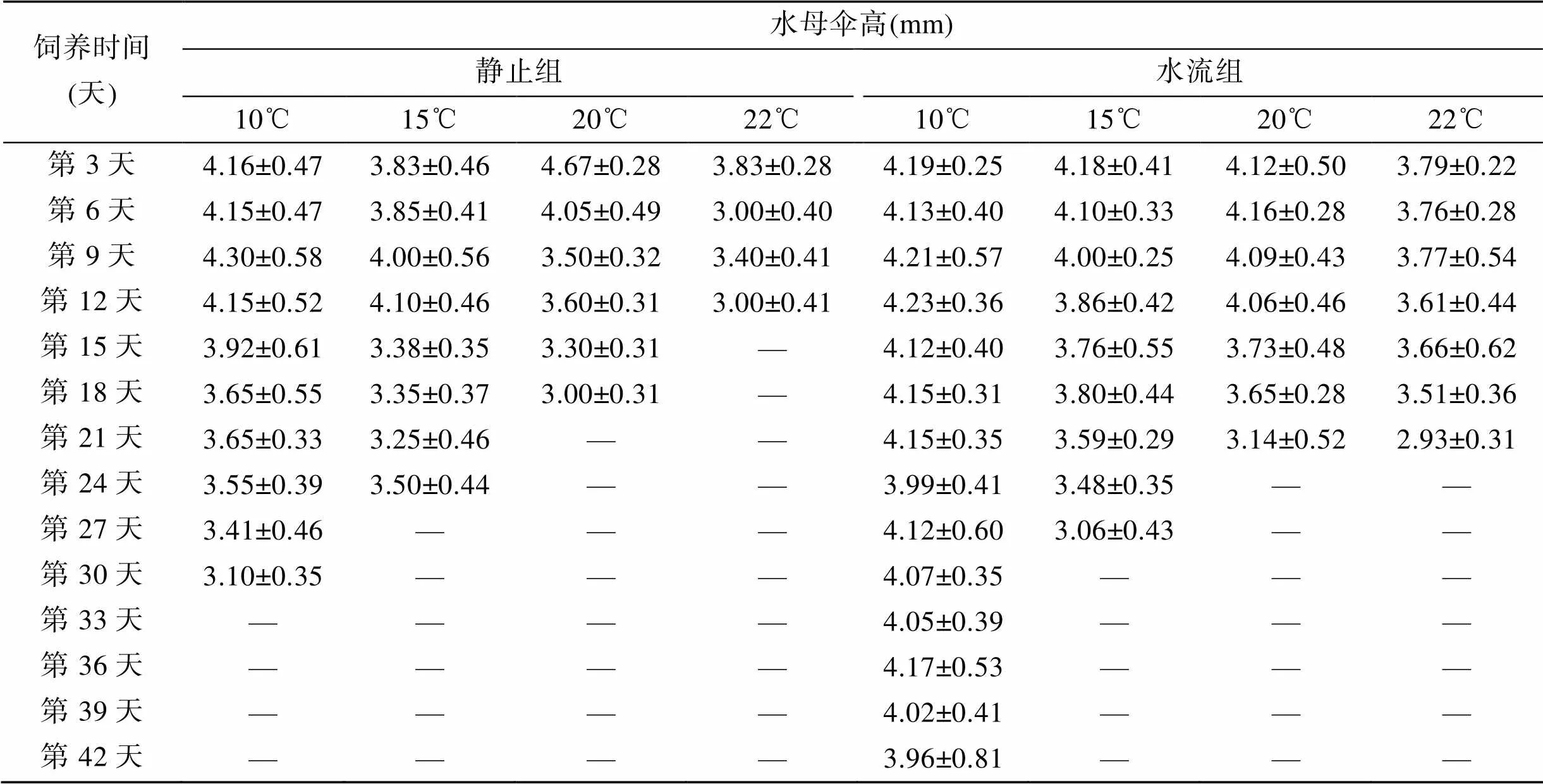
表2 不同条件下水母伞高

续表
试验用八斑唇腕水母伞径︰伞高为1︰1.12, 经人工饲养后, 伞径: 伞高为1︰1.13, 伞径和伞高相关系数=0.95, 说明在饲养过程中, 随着伞径的缩小, 伞高也在萎缩, 两者密切相关。
经Duncan法进行多重比较, 静止组10℃与15℃饲养条件下水母的伞径和伞高差异不显著(>0.05), 与其他组差异均极显著(<0.01)。水流10℃组水母的伞径和伞高与其他组差异均极显著(<0.01)。无论是否有水流, 20℃和22℃组间差异均不显著(>0.05)。说明10℃是人工养殖采集的八斑唇腕水母的最佳条件, 15℃短期内也能满足该水母的生存需要, 但是更高的温度将会引起水母死亡。
10℃饲养条件下, 水流组水母伞径和伞高极显著大于静止组(<0.01)。而其他温度组间差异不显著(>0.05)。说明温度是影响八斑唇腕水母生存的决定因素, 在低温条件下, 水流对该水母的长期生存也是必须的。温度越高, 水母越容易萎缩, 而水流的影响越小。
2.2 温度和水流对分裂数影响
表3显示了不同条件下30只水母的分裂数。静止10℃、15℃、20℃和22℃组在饲养3 d内分别分裂1、3、4和5只水母幼体, 除了10℃组在以后饲养的18 d正常分裂外, 其他3组3 d后很少分裂。水流10℃组水母在57 d内保持形态正常, 同时正常产生水母芽, 其他温度下产生水母幼体的趋势与静止烧杯中饲养的水母相似, 即随着人工饲养时间的延长, 每天产生的水母幼体数量越来越少, 并且相同的水流条件下, 温度越高, 最初产生的水母幼体数量越多, 但是持续时间越短。无论是否存在水流, 试验水母在萎缩死亡前6 d不再分裂幼体。10℃组在水流静止的条件下30 d分裂16个水母幼体, 在77 cm/s水流的环境中能存活64 d, 分裂25个幼体, 水母出芽率分别为53.3%和83.3%。其次为15℃水流组, 在存活的27 d内分裂11个水母幼体, 而分裂数量最少的均出现在水流静止组, 15℃和20℃组分别分裂3个和4个幼体。
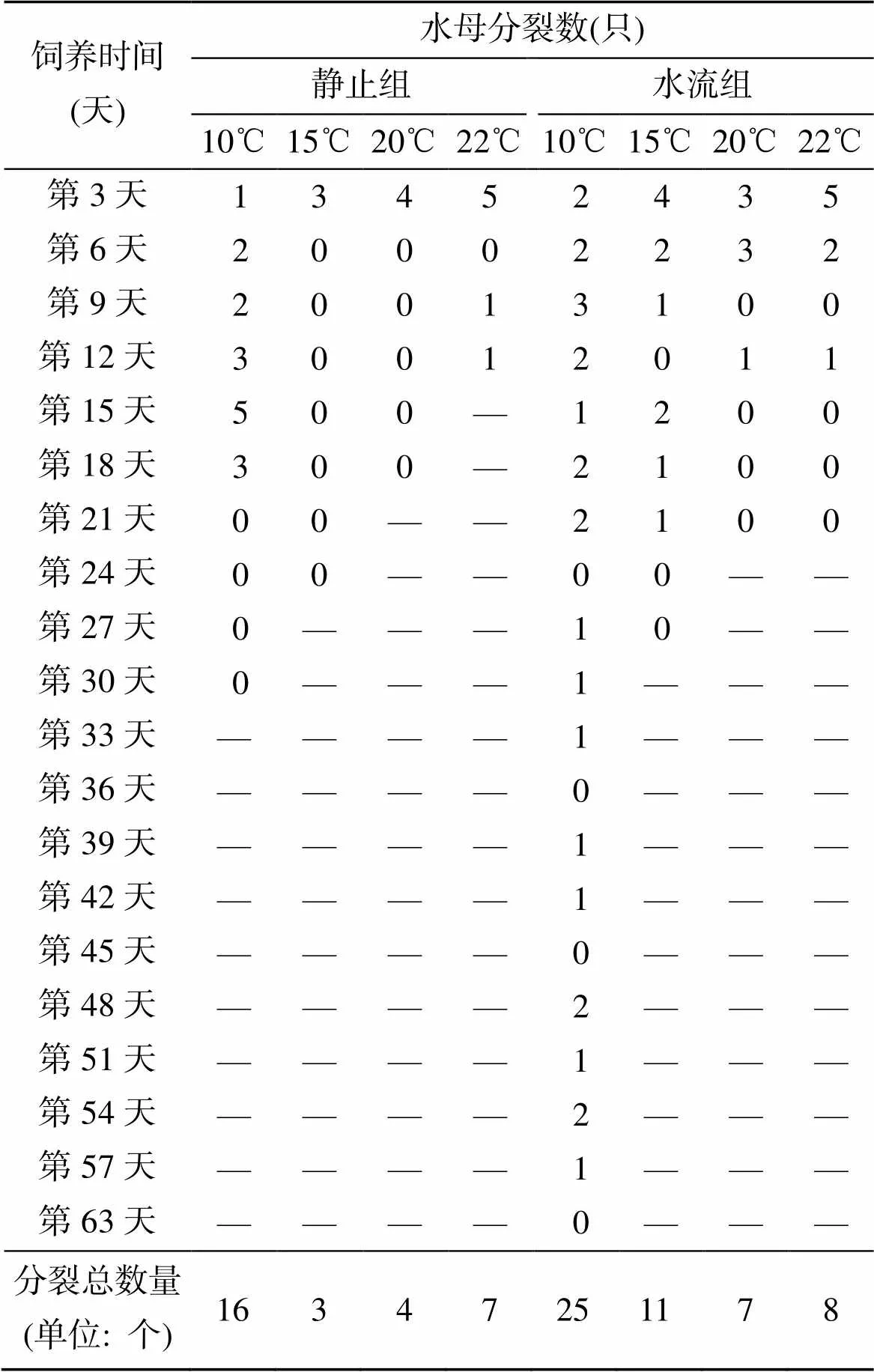
表3 不同条件下水母分裂数
2.3 温度和水流对存活时间的影响
从图4、5可以看出, 温度是影响该水母存活的首要因素, 温度越低, 水母存活时间越长。无论是否存在水流, 10℃组水母的存活时间均最长, 分别为33和64 d。而20℃和22℃组均在饲养9 d内, 部分成体水母开始死亡, 平均存活时间小于16 d。

图4 静止组水母存活情况
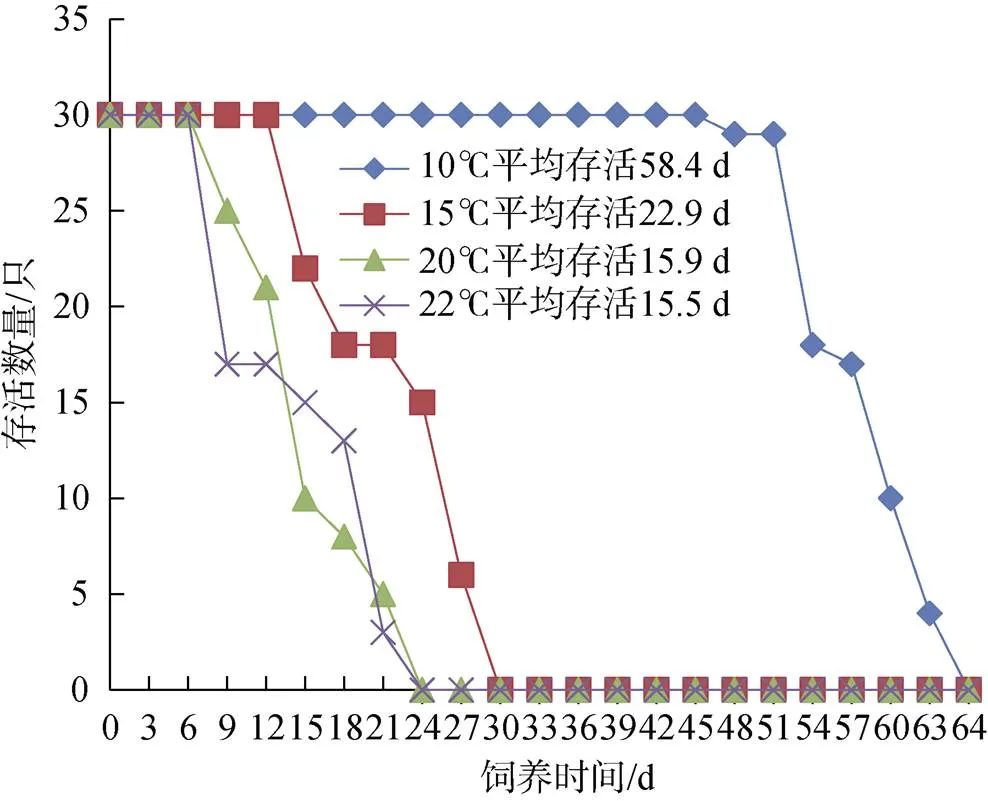
图5 水流组水母存活情况
水流对20℃和22℃组水母存活时间影响较小, 但合适水流将10℃组水母平均存活时间从19.9 d延长到58.4 d, 也将15℃组水母的平均存活时间延长了4.8 d。静止10℃组较15℃组水母平均存活时间仅长1.8 d, 在饲养第18天时, 15℃组较10℃组多存活2只水母, 说明在静止水流中, 两种温度对该水母影响差异很小。但在合适水流条件下, 10℃组水母最长能存活64 d, 而15℃组仅存活30 d, 说明水流与温度共同作用, 是水母正常生存的必须要素。
3 讨论
3.1 温度影响八斑唇腕水母的形态和丰度
八斑唇腕水母在胶州湾是冬季的优势种类。2003年胶州湾浮游生物浅水I型网中水母的调查显示, 八斑唇腕水母2、3月份的丰度均很高, 分别为64.7 个/m3、118.0个/m3, 分别占水母总平均丰度的99.5%和98.0%[11]。但在其他季节, 随着水温升高, 该水母的丰度变少, 除2006年4月石岛外站占绝对优势外, 其余7调查航次丰度很小或者未发现[12]。在低温低盐条件下, 八斑唇腕水母采食水中浮游动物快速生长繁殖的同时, 也可以作为其他浮游动物的饵料, 黄有松等[13]以核糖体小亚基基因(18S rDNA)为目标序列, 研究了青岛近海中华哲水蚤的现场食物组成, 其中含有八斑唇腕水母基因序列的样品占总样品的40%以上。本试验每天投喂1次孵化24 h的卤虫无节幼体, 水母采食正常, 水流10℃组57 d仍能分裂水母幼体, 成体水母在人工条件下可以存活64 d, 说明饵料和水流能满足水母生长发育要求。
钵水母纲水母(Scyphozoa)生活史研究较多, 咖啡金黄水母()的碟状体在合适的水流和充足的食物条件下, 60 d内可以变态为伞径10 cm的水母育成体, 水母成体能在人工条件下饲养8~10个月[14]。水螅纲水母体的存活时间较钵水母纲短, 仅为60 d~90 d。王丹丽等[15]在不同温度、盐度、pH等胁迫条件下, 研究了信阳桃花水母的存活情况, 结果表明, 在合适饲养条件下, 水母在试验结束时, 形态保持正常, 存活率100%, 并且桃花水母的耐高(低)温能力与其生活状态密切相关。本试验在水流10℃组八班唇腕水母伞径、伞高能长时间保持正常, 并且水母持续性出芽产生幼体, 存活时间达到64 d, 与其他水螅类水母生命期接近, 说明八斑唇腕水母可以在10℃、合适的水流条件下人工繁育。
3.2 合适的水流促进水母发育和存活
在自然界中, 水流为水母带来食物和新鲜的水质。在人工饲养过程中, 水流速度和方向也是水母繁育的关键因素。与钵水母具有纵肌和环肌不同, 水螅水母依靠伞缘的缘膜和外伞表面发育程度低的上皮细胞, 借助水的动力游泳。本试验在封闭的烧杯和循环缸内进行, 充气带动水流的循环缸内水质稳定均一, 并且水流可以帮助水母自由游动。而在没有水流情况下, 水螅水母游动能力弱, 采食前可以悬浮在水中, 但采食后主要沉积在容器底部。本次试验结果表明, 合适的水流促进了水母幼体的分裂, 延长了水母的寿命, 是人工饲养八斑唇腕水母的必要条件。
刺胞动物生活史由水螅型世代和水母型世代组成, 水螅型世代用无性出芽生殖方式产生水母型世代, 而水母型个体脱离母体, 长大成熟以后, 又以有性生殖方式产生水螅型个体, 两个世代相互交替进行, 仅有少数种类缺少某个阶段。与其他刺胞动物的水母体从水螅体或者浮浪幼虫发育不同, 八斑唇腕水母体既可以从水螅体无性出芽生殖, 又可以从成熟的水母体出芽发育而来, 在科学研究和科普展示等方面都具有特殊性和代表性, 值得长期饲养展示。鉴于该水母个体较小, 伞径仅为3.65 mm, 经管道水流冲击变形后, 能通过40目的隔栅网, 建议在饲养工作中, 采用换水量小的滴水水浴缸或者完全封闭的充气水浴缸饲养展示。
3.3 水温促进生殖腺的成熟
水温是影响动物生殖腺发育的主要外界因素, 生物学上将某种生物的生殖腺必需在外界水温达到某一特定的下限水温称为生物学零度, 而要使生殖腺发育成熟, 必需在其生物学零度以上经一段时光的积聚, 即有效积温。如海湾扇贝在水温17℃, 积温267℃时, 性腺达到成熟期, 水温21℃, 积温446℃时, 性细胞开始排放[16]。Lucas[17]认为, 温度通过直接调节海月水母的代谢速率及间接提高水体的浮游动植物数量, 使海月水母快速生长, 并在个体较小的情况下达到性成熟。本试验中, 静止和水流条件下, 22℃组虽然存活时间仅为15和24 d, 但第3天分裂幼体均为5个, 而10℃组分别为1个和2个, 可能由于高温促进了水母细胞的增长和分化, 导致生殖腺的快速成熟。
3.4 野生水母的人工驯化和人工繁育
八斑唇腕水母是偏低温低盐的种类, 广泛分布于北大西洋沿岸、北太平洋沿岸、黑海、地中海和北极海等边缘海域[18]。在中国出现在冬、春季的黄海及其以北海域, 以中国内海——渤海最多, 浙江和福建沿海也有少量分布[19]。与大型水母经常暴发并对渔业资源和渔业生产造成较大影响不同[20], 目前水螅水母的丰度变化与海洋生态和经济关系的研究较少。本次试验材料采集于黄海西北部, 属温带季风气候, 2016年3月10次共采集914只八斑唇腕水母, 这与该区域水温稳定、风力小、饵料丰富等环境条件密切相关。
野生鱼的人工饲养和繁育已被深入研究[21], 但水母的相关报道很少。由于水母存在世代交替, 在自然界中具有明显的季节性, 为保证长期饲养研究, 水母在人工条件下的驯化和繁育更加重要。本试验水母采集温度9.5℃~14℃, 最好的人工饲养水温也应该是9.5℃~14℃。但是如果作为科普展示种类, 满足此条件对设备要求很高, 因此希望在保证水母展示效果和存活时间的前提下, 找到一个合适的温度饲养该水母。本试验结果表明, 温度越低, 水母生长越好, 存活时间越长, 水流对水母的正常生长发育是必需的。在合适水流情况下, 10℃能保持水母自然形态和存活。但由于八斑唇腕水母具有特殊的水母出芽生殖, 伴随着水母幼体的不断产生, 在实际饲养工作中, 当温度短期内升高到15℃时, 水母也能依靠自身的繁育持续存活。
[1] 许振祖, 黄加祺, 林茂, 等. 中国刺胞动物门水螅虫总纲[M]. 北京: 海洋出版社, 2014, 267-269. Xu Zhenzu, Huang Jiaqi, Lin Mao, et al. The superclass Hydrozoa of the Phylum Cnidaria in China[M]. Beijing: China Ocean Press, 2014, 267-269.
[2] Russell F S. Hydromedusae. Order: anthomedusae, families: rathkeidae and bougainvilliidae[J]. Zooplankton, 1953, 51: 1-4.
[3] Werner B. Die Verbreitung und das jahreszeitliche auftreten der anthomeduseM. Sars, sowie die temperaturabhängigkeit ihrer entwicklung und fortpflanzung[J]. Helgoländer Wissenschaftliche Meeresuntersuchungen, 1958, 6(2): 137-170.
[4] Berrill N J. Development and medusa-bud formation in the hydromedusae[J]. The Quarterly Review of Biology, 1950, 25(3): 292-316.
[5] Bouillon J, Bernhard W. Production of medusae buds by the polyps of(M.Sars) (Hydroida Athecata)[J]. Helgoländer Wissenschaftliche Meeresuntersuchungen, 1965, 12(1/2): 137-148.
[6] Russell F S. The medusae of the British Isles II[M]. Cambridge: Cambridge University Press, 1970: 284.
[7] 陈玉龙, 王旭, 胡锦矗. 桃花水母生活史的实验观察[J]. 动物学杂志, 2010, 45(2): 88-92. Chen Yulong, Wang Xu, Hu Jinchu. Laboratory observations on the life cycle of[J]. Chinese Journal of Zoology, 2010, 45(2): 88-92.
[8] O’Sullivan D. Description ofsp. nov. (Hydromedusae: Rathkeidae) from Antarctica and a key to the genus[J]. Journal of Natural History, 1984, 18(6): 861-868.
[9] Wang C G, Xu Z Z, Huang J Q, et al. Taxonomic notes on Hydroidomedusae (Cnidaria) from South China Sea Ⅲ: Family rathkeidae and zancleopsidae[J]. Zoological Systematics, 2016, 41(4): 392-403.
[10] 黄苑媚, 刘志刚, 谢恩义, 等. 水流速率对全缘马尾藻幼孢子体生长和生理活性的影响[J]. 广东海洋大学学报, 2014, 34(6): 45-50. Huang Yuanmei, Liu Zhigang, Xie Enyi, et al. The effects of water velocity on the growth and physiological activities of juvenile sporophyte of[J]. Journal of Guangdong Ocean University, 2014, 34(6): 45-50.
[11] 张芳, 杨波, 张光涛. 胶州湾水母类生态研究Ⅱ. 优势种丰度的时空分布[J]. 海洋与湖沼, 2005, 36(6): 518-526. Zhang Fang, Yang Bo, Zhang Guangtao. Ecology of medusa in Jiaozhou Bay II. Spatial and temporal distribution of dominant species[J]. Oceanologia et Limnologia Sinica, 2005, 36(6): 518-526.
[12] 孙松, 张芳, 李超伦, 等. 黄海小型水母的分布特征[J]. 海洋与湖沼, 2012, 43(3): 429-437. Sun Song, Zhang Fang, Li Chaolun, et al. The distribution pattern of small medusa in the Yellow Sea[J]. Oceanologia et Limnologia Sinica, 2012, 43(3): 429-437.
[13] 黄有松, 徐东晖, 陈洪举, 等. 黄河口邻近水域中华哲水蚤现场食物组成的分子生物学检测[J]. 中国海洋大学学报(自然科学版), 2014, 44(3): 83-91. Huang Yousong, Xu Donghui, Chen Hongju, et al. PCR-Baseddietary analysis ofBrodsky in Yellow River Estuary and adjacent waters[J]. Periodical of Ocean University of China, 2014, 44(3): 83-91.
[14] 杨翠华, 王玮, 王文章, 等. 咖啡金黄水母的人工培育及生活史观察[J]. 水产科学, 2012, 31(12): 708-712. Yang Cuihua, Wang Wei, Wang Wenzhang, et al. Laboratory observations on the life-cycle of the brown sea nettle jellyfish[J]. Fisheries Science, 2012, 31(12): 708-712.
[15] 王丹丽, 徐善良, 蒋宏雷, 等. 信阳桃花水母对几种生态因子胁迫的耐受反应[J]. 应用生态学报, 2006, 17(6): 1103-1106. Wang Danli, Xu Shanliang, Jiang Honglei, et al. Tolerance ofto the stresses of some ecological factors[J]. Chinese Journal of Applied Ecology, 2006, 17(6): 1103-1106.
[16] 吕豪, 李霞, 董义超, 等. 海湾扇贝种贝人工促熟条件下积温和性腺发育周期的关系[J]. 海洋科学, 2008, 32(4): 57-60. Lv Hao, Li Xia, Dong Yichao, et al. The relation between effective accumulated temperature and gonad development period of scallop,[J]. Marine Sciences, 2008, 32(4): 57-60.
[17] Lucas C H, Lawes S. Sexual reproduction of the scyphomedusain relation to temperature and variable food supply[J]. Marine Biology, 1998, 131: 629- 638.
[18] Kramp P L. Synopsis of the Medusae of the world[J]. Journal of the Marine Biological Association of the UK, 1961, 40: 7-469.
[19] 张金标. 中国海域水螅水母类区系的初步分析[J]. 海洋学报, 1979, 1(1): 127-137. Zhang Jinbiao. A preliminary analysis on the Hydromedusae fauna of the China sea areas[J]. Acta Oceanologia Sinica, 1979, 1(1): 127-137.
[20] 张芳, 李超伦, 孙松, 等. 水母灾害的形成机理、监测预测及防控技术研究进展[J]. 海洋与湖沼, 2017, 48(6): 1187-1195. Zhang Fang, Li Chaolun, Sun Song, et al. Progress on studying jellyfish bloom, and the monitoring and control[J]. Oceanologia et Limnologia Sinica, 2017, 48(6): 1187-1195.
[21] 林越赳, 蔡良候, 曾庆民, 等. 台湾海峡野生牙鲆人工育苗技术研究[J]. 海洋科学, 2006, 30(2): 91-93. Lin Yuejiu, Cai Lianghou, Zeng Qingmin, et al. Study on the breeding ofdistribution in the Taiwan Straits[J]. Marine Sciences, 2006, 30(2): 91-93.
Effects of water temperature and velocity on the development of
YANG Cui-hua, WANG Wen-zhang, WANG Hai-ming
(Marine Science and Technology Museum, Qingdao 266003, China)
is a smalljellyfish that is widely distributed in the North Atlantic coast, the North Pacific coast, the Black Sea, the Mediterranean Sea, the Arctic and other marginal waters. In recent years, this species has gained more attention because of its unique reproduction cycle. To enable further study, a large number of jellyfish have been kept indoors for a long time, and a life cycle pattern has been observed. Therefore, to optimize artificial breeding and facilitate scientific study of this species, we examine the effects of water temperature and velocity on the diameter, height, budding, and survival time of these jellyfish. Lots of jellyfish were collected along the coast of Lu Xun Park in March 2016 with the surface water temperature 9.5℃- 14℃. After 24 h of temporary cultivation,were raised under the cross conditions of different water flow rates (1 L beaker with stationary water, 13 L tank with bubbles to drive water) and different temperatures (10℃, 15℃, 20℃, or 22℃). The results showed that the survival time in 10℃ group was the longest when the water velocity was 77 cm/s. However, this was negatively related to the umbrella diameter, umbrella height, and feeding time. Higher temperatures proportionally reduced the survival time of the jellyfish. Temperature promoted rapid maturation of gonads, but budding was persistent in the 10℃ group. The survival time was more 38.5 d under the water flow 77cm/s than the static environment at 10℃ group. The study shows that water velocity promoted the activity of jellyfish and significantly prolonged survival time.can be bred through polyp and gonadal reproduction, and it preferentially grows at low temperatures in water baths. These data may help to promote and enhance the captive breeding and display of these remarkable jellyfish for research and educational purposes.
; temperature; water velocity; budding; survival time
Jun. 6, 2018
[Science and Technology Development Project of Qingdao Shinan District, No.2018-4-011-ZH]
Q959.132.2
A
1000-3096(2020)04-0067-08
10.11759/hykx20180606002
2018-06-06;
2019-06-01
青岛市南区科技发展资金项目(2018-4-011-ZH)
杨翠华(1978-), 女, 山东寿光人, 高级工程师, 博士, 主要从事观赏性海洋动物繁育研究, 电话: 0532-82890112, E-mail: ycuihua@hotmail.com
(本文编辑: 谭雪静)
