模拟增温对水葱(Scirpus validus)输导组织的影响1)
2020-05-12冯春慧孙梅田昆管东旭张依南
冯春慧 孙梅 田昆 管东旭 张依南
(国家高原湿地研究中心(西南林业大学),昆明,650224)
植物的维管结构(plant vascular system)是植物体中担负长距离物质运输的主要输导结构,主导着植物体各部分间的物质重新分配和转移,关系着植物的整个生命过程[1-2]。维管结构主要由木质部和韧皮部组成[3],其中,导管是木质部最主要的组成部分,导管孔径的大小和密度直接影响水分运输效率[4-5]。而韧皮部的筛管是长距离运输光合产物的重要结构,且物质的运输效率与筛管密度和大小密切相关[6]。
温度是影响植物生长发育的关键因素之一,输导组织以及光合作用对温度变化的响应与植物的水分传输和利用、物质积累等生理过程密切相关,对了解植物如何适应温度变化有重要作用。相对于低温环境,生长在较高温度下的烟草叶片具有更大的导管直径并对应更高的导水率[7]。对生长在不同温度条件下早熟禾的研究发现,生长在较温暖条件下的早熟禾具有更大和更密的导管,以提高植物水分输送能力[8]。但也有少部分研究认为,增温对植物的输导组织影响不显著[9]。植物光合功能受环境温度的影响也十分显著,温度通过影响植物对光和CO2的利用效率影响植物的光合作用,进而影响植物的物质积累和生长繁殖[10]。但是不同植物对增温会产生不同的光合生理响应[11-12]。例如,增温使单子叶草本植物发草(Deschampsiacaespitosa)的净光合速率、气孔导度、蒸腾速率显著提高;但增温条件下的双子叶草本植物遏蓝菜(Thlaspiarvense)的净光合速率、气孔导度和蒸腾速率都明显低于对照组[13]。前期我们对滇西北高原纳帕海湖滨带典型优势植物适应增温的研究也发现,增温显著降低了茭草的光合功能,而对黑三棱的影响相对较弱[12]。也有研究表明,增温降低了芦苇的净光合速率、蒸腾速率和气孔导度[14]。植物对增温的反应存在一定的有效幅度,适度增温能使植物的净光合速率和蒸腾速率等光合特性指标增加,但当环境温度超过植物最适生长温度范围时,增温将会对植物的光合作用产生抑制作用[15-16]。可见,增温对植物维管组织及光合功能的影响尚存在众多争议,仍需进一步在各类型植物中开展相关研究。此外,在陆生植物类群之中,植物净光合速率与其水分运输能力间存在的正相关关系已被诸多研究所证实[17-18]。植物输导组织与光合功能间密切的协同作用是植物实现对增温的响应与适应的重要策略之一[19]。目前对温度变化如何影响植物输导组织和光合功能的研究大多集中于森林生态系统,研究对象多集中于木本植物,而对其他生态系统和植物类群的研究相对较少。
湿地生态系统是近年来各国研究的热点,湿地植物作为湿地生态系统中的重要生产者,对整个湿地生态系统的构建、维持和恢复具有重要意义[20-21]。研究湿地植物对温度变化的响应规律是掌握气候变化对湿地生态系统影响的基础。云南高原湿地是我国高原湿地的重要组成部分,由于地处较高海拔,地形相对封闭的特征,对气候变化高度敏感,更容易受到全球增温效应的影响[22]。水葱(Scirpusvalidus)的分布范围较广,是典型的湿地植物,在滇西北著名的九大高原湖泊中都是其湖滨带优势植物[23]。前期对滇西北水葱光合生理的研究发现,水葱的光合功能性状对温度变化较为敏感[22]。因此,水葱在研究增温对湿地植物应对气候变化的适应策略方面是较为理想的材料。
本研究以云南高原典型的湿地植物水葱(Scirpusvalidus)为研究对象。基于IPCC对未来大气增温的预测,采用闭合式人工控制气候模拟实验系统开展增温控制实验,通过观测不同温度处理下水葱的叶片输导组织及净光合速率变化情况,探讨植物的输导组织和光合生理功能对增温的响应和适应策略,为揭示高原湿地植物对气候变暖的生理适应策略提供案例。
1 研究地概况
本研究实验材料移栽自云南省晋宁县滇池生态定位站附近的滇池湖滨带(102°35′54″~102°40′8″E,24°39′59″~24°42′17″N),海拔1 888 m。滇池(24°22′~26°33′N,102°10′~103°40′E),地处云南省昆明市西南部,平均海拔1 943 m。拥有盘龙江等21条补给河流,流域面积(不包括海口以下流域面积)2 960 km2,平均水深4.5 m。流域范围属北亚热带湿润季风气候,干湿季分明。全年温差较小,年平均气温15.1 ℃,年均日照2 200 h;霜期短,无霜期240 d以上;年降水量1 035 mm。鸟类、鱼类和湿地植物资源十分丰富,如中山杉(Ascendensmucronatum)、芦苇(Phragmitescommunis)、水葱(Scirpusvalidus)、香蒲(Typhaorientalis)等,整个湿地的植被类型以乔木和灌木、沉浮水植物群落类型为主,总面积占整个湿地总面积的72.95%[24]。
2 材料与方法
2.1 环境模拟控制实验系统
基于IPCC第五次评估报告,在云南省昆明市国家高原湿地研究中心建立人工环境控制系统封顶式生长室(图1),设置对照、大气增温(环境温度+2 ℃)两个不同处理。生长室下部近似圆柱体,由11个宽1.0 m和高2.0 m的面构成,用浮法玻璃密封;上部近似球缺,高约1.5 m,由双层中空PC板密封,总体积约为24.5 m3。每个生长室具有独立、自控的特点。增温生长室的下部圆柱体一边侧面与制冷-加热系统连接,生长室内温度的控制主要是通过压缩机的制冷功能和与其相连接的热交换器、电阻加热器以及新风量控制阀等控制,为了维持生长室内气压平衡和避免特殊事件(如停电、空调故障)造成生长室内温度升高,在球缺顶部装有2个自垂式单向开启调压扇和1个手动窗口。

图1 人工环境控制系统封顶式生长室(STC)
2.2 人工控制气候模拟实验生长室温度变化
生长室的运行状态和数据采集是由模拟控制监控系统实现,该系统通过通讯线监视各个封顶式生长室的当前运行状态,并可远程设定系统的运行参数以及进行系统环境数据的不间断采集。用Excel统计水葱生长季(3—10月)生长室内的温度数据,并分别计算日平均温度和月平均温度的平均值。根据这些温度值,相比于对照组,增温生长室内的大气平均温度在试验期间(2017年3月—2017年9月)增加了1.96 ℃(图2)。这一结果说明闭合式人工模拟温度控制系统实现了显著增温作用,且增温幅度和在预测范围内,模拟增温效果比较理想。
2.3 实验条件设置
在滇池湖滨带选择长势基本相似,生长良好的水葱植株,带土采集并移至气候模拟实验基地。将这些植株移栽到8个内径35 cm,高25 cm的实验桶内,每个实验桶内的栽培基质均为水葱采样点的原位土壤,每桶放入相同的土量。在自然条件下缓苗半个月后,将8盆水葱随机分成两组(每组4盆),一组放入现行环境温度的生长室,作为对照组;将其余4盆放入温度设定为现行环境温度+2 ℃的生长室。为了保持均一的光照条件,排除边缘效应,放置时以生长室的中心为圆点,在距生长室内边缘至圆心的中间位置随机放置。让水葱在生长室内生长1年(2016年10月—2017年10月),以便水葱充分适应增温环境。在实验期间,保持每个桶里的水葱淹水深度一致。

CK=环境温度,TE=(2±0.5)℃增温。
2.4 净光合速率的测定
于2017年8月,选择晴朗天气,于上午08:30—11:30,用Li-6400XT便携式光合作用测定仪(LI-6400,LI-COR,Nebraska,USA),分别从不同处理的4盆水葱中各挑选3株长势一致、成熟并且无损伤的水葱,每株选择一片的全展成熟叶片,原位直接测定植物的净光合速率(Pn)。测定时,设定叶室内部光强为1 500 μmol·m-2·s-1,叶片温度保持在22~24 ℃,流速设为500 μmol·s-1,室内CO2摩尔分数设置为400 μmol·mol-1。
2.5 输导组织性状测定
将测定完净光合速率的叶片截取中间部位长度约15 cm的叶片,密封于含湿纸球的密封袋内,做好标记并保存于保鲜盒内,带回实验室进行维管组织的解剖及测定。在实验室中,分别对截取的叶片进行徒手切片,在光学显微镜下观察并拍照。然后用Image J图片处理软件测量和统计各输导组织的导管横切面长度、宽度和导管横切面面积以及筛管横切面的密度和面积,并以椭圆的周长计算公式算出导管横切面的周长(图3)。每个处理选3株植物,每株植物选择8个维管束结构。即每个温度处理下的每个导管和筛管的参数值各24个数值,总共48个值。
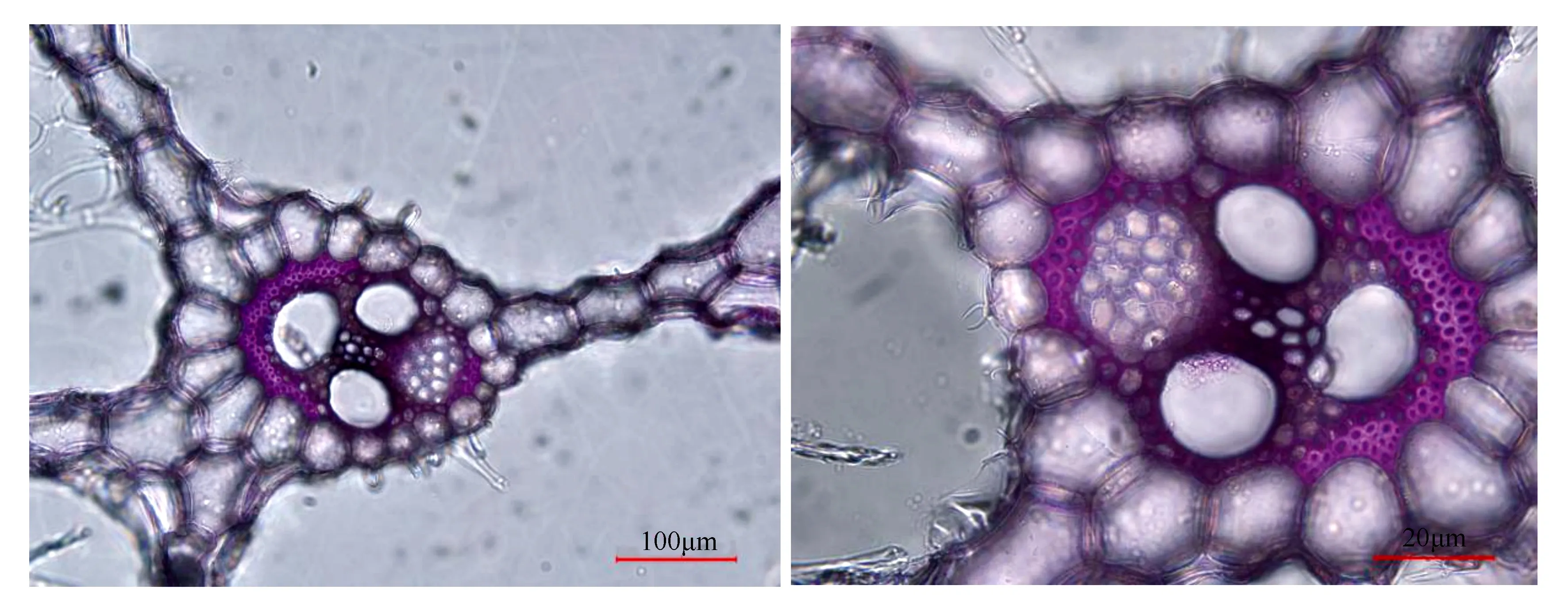
图3 水葱维管束解剖结构(左图为放大20倍镜像,右图为放大40倍镜像)
2.6 数据分析
本研究采用SPSS统计分析软件,并通过独立样本t-检验检测不同温度处理下水葱的输导组织及光合性状间有无显著差异,统计的显著性水平为P<0.05。采用R统计分析软件进行线性相关分析,检测水葱输导组织性状与净光合速率、气孔导度和蒸腾速率间的相关关系。采用Sigmaplot(v.10.0;https://systatsoftware.com/)绘图软件进行绘图。
3 结果与分析
3.1 水葱的输导组织和净光合速率在不同温度处理间的差异
相对于对照组,增温处理下的水葱导管横切面周长、导管横切面面积、筛管横切面面积和净光合速率(Pn)均呈显著减小的趋势(P<0.05,表1),其平均值分别减小了7.2%、10%、14.7%、47.5%。但是增温处理下筛管密度显著高于对照组(P<0.05),其平均值高于对照组20.2%。
2.2 植物维管束结构与净光合速率的相关性
基于对水葱的维管结构性状与净光合速率的相关分析表明:水葱的导管横切面周长、导管横切面面积与净光合速率间都存在显著正相关(P≤0.05),而筛管横切面密度与净光合速率间呈现显著负相关(P<0.05),筛管横切面面积与净光合速率间关系不显著(图4)。

表1 水葱的输导组织和净光合速率在不同处理间的差异比较
注:表中数据为平均值±标准差;同列不同字母代表差异性显著(P<0.05)。

r为水葱净光合速率与维管结构性状之间的相关系数;P为显著性水平;P≤0.05为显著相关;P>0.05不相关。
3 结论与讨论
增温显著降低了水葱的导管横切面周长、导管横切面面积、筛管横切面面积和净光合速率,但显著增加了筛管横切面密度。这一结果表明,增温显著抑制了水葱输导组织以大小为基准的水分和营养传输能力以及光合生产力。水葱的净光合速率与导管横切面周长、导管横切面面积和筛管横切面面积均呈正相关,但与筛管横切面密度呈负相关,表明水葱的净光合速率与其输导组织结构密切相关。
输导组织作为植物体内物质运输的重要组成部分,其解剖结构差异反映了植物对外界环境改变所产生的生理适应策略。植物的导管和筛管的大小和数量是影响植物体内水分和养分运输的重要因素。一般来说,导管和筛管的数量越多、面积越大则传输效率越高[25]。对木本植物的研究发现,温度升高会加大植物的水分胁迫[26],木本植物应对水分胁迫时往往会增大导管大小以及密度来提高水分的运输能力[27]。而本研究结果发现,增温反而减小了水葱的导管横切面大小,且水葱每个维管束内导管数量都仅有两个,因此,增温并没有增加水葱的水分运输能力,相反是降低了。这可能是由于水葱是水生植物,水分并不是水葱的生长限制因子,因而受增温引起的水分胁迫影响较小。此外,温度增高导致水葱蒸腾速率加大,水分散失较快,极易引起植物生理水分亏缺[28-30]。而较小的导管直径可以增加水分的运输阻力[31],防止体内水分的过度散失。因此,水葱的导管横截面周长和面积变小可能是面对高温情况下,其生理结构的一种保水策略。
过去关于增温对植物净光合速率的影响存在众多争议,本研究结果发现,增温显著降低了水葱的净光合速率(P=0.002),这与祁秋艳等[14]对崇明东滩芦苇的研究和陈翔等[32]对高寒草甸植被的研究结果一致。水葱长期生活在滇池湖滨带,暗示滇池自然界的环境温度是其最适生长的温度,增温实验导致其生存的平均温度、最高气温和生长季积温都明显增加,很可能引起水葱对较高温度的不适应,产生高温胁迫反应。高温胁迫会使光合作用的关键酶Rubisco活化酶失活[33],RuBP羧化酶活性降低[34],致使光合速率显著下降。
植物的光合作用往往和植物的水分吸收、运输和蒸腾过程密切相关[19]。在陆生植物类群之中,植物的净光合速率和其水分运输的能力之间存在正相关关系已经被证实[17-18]。本研究结果发现,水葱的净光合速率与其导管横截面大小显著正相关,说明水葱的净光合速率与水分运输能力存在正相关。植物通过体内的导管和筛管等输导组织结构运输水分和营养物质,供其进行光合作用和呼吸作用,其光合产物又通过筛管往回运输,供植物生长和繁殖。筛管是植物体内光合产物的重要运输通道。水葱筛管横切面大小与净光合速率的正相关,可能是因为植物的净光合速率下降,光合产物积累也减少,筛管的运输量下降而导致的筛管横切面面积变小。
增温显著降低了水葱的水分和物质运输能力和光合能力,可能将进一步引起植物的生长和扩散受到抑制。由于水葱是云南大部分高原湿地的优势植物,增温对水葱输导组织结构和光合速率的影响,反映了这一高原地区湿地植物对气候变化的适应策略。据此推测,未来气候变暖可能会对云南高原湖滨带植物的结构组成产生深远的影响。本研究结果可为高原湖滨带植物应对气候变化的适应性研究提供案例。
