雷帕霉素联合甲霜·噁霉灵对马铃薯早疫病菌的抑制效应
2020-05-06吕秀兰金良郑先喆张美权董攀李正国任茂智
吕秀兰 金良 郑先喆 张美权 董攀 李正国 任茂智

摘 要 2018年3—5月,利用菌丝生长速率法,测定雷帕霉素对马铃薯早疫病的抑菌活性,雷帕霉素联合甲霜·噁霉灵的协同效应;并通过透射电镜及扫描电镜在超微结构上观察,揭示雷帕霉素抑制马铃薯早疫病的机制。结果表明,雷帕霉素能有效抑制病原菌菌丝生长;雷帕霉素可导致病原菌菌丝节间膨大,细胞隔膜消失,胞内细胞器消失及细胞死亡;雷帕霉素联合甲霜·噁霉灵具有明显的协同效应。甲霜·噁霉灵单独处理时,其IC50药物浓度约为0.5 mL·L-1;雷帕霉素+甲霜·噁霉灵组合处理时,达到IC50所需要的甲霜·噁霉灵的浓度降低为0.05 mL·L-1。
关键词 雷帕霉素;甲霜·噁霉灵;马铃薯早疫病菌;抑菌效果;协同作用
中图分类号:S436.32 文献标志码:A DOI:10.19415/j.cnki.1673-890x.2020.7.001
马铃薯是世界第三大粮食作物[1]。马铃薯早疫病在世界各大马铃薯种植区域均有发生,被普遍认为是继晚疫病后影響马铃薯生产的第二大病害[2-3]。马铃薯早疫病的病原菌主要有茄链格孢菌(Alternaria solani)、链格孢菌(Alternaria alternate)、极细链格孢菌(Alternaria tenuissima)等[4],其中茄链格孢菌是优势病原菌,归属于半知菌纲链格孢属。由于化学农药防控具有快速、防效稳定和操作简便等优点,目前仍是防治马铃薯早疫病的主要方法[5]。但是长期大量使用化学农药会导致环境污染、危及人体健康及食品安全等严重问题,因此,研发有效、安全及环保的生物农药是保障马铃薯生产,减少环境污染的重要措施之一。
雷帕霉素(Rapamycin,RAP)又名西罗莫司(Sirolimus),是一种大环内酯类免疫抑制剂,主要作用于TOR蛋白激酶(Target of Rapamycin)。TOR蛋白激酶具有高度的保守性,普遍存在于真核细胞中,其突变可诱使细胞死亡[6]。雷帕霉素能与FKBP12蛋白结合形成二元复合物[7],该复合物再与TOR蛋白激酶的FRB结构域结合,从而抑制TOR蛋白激酶的活性。已有研究表明,雷帕霉素对番茄灰霉病菌、油菜菌核病菌及棉花枯萎病菌等表现出极强的抑制活性[8]。但其是否能够有效抑制马铃薯早疫病菌及其与农药混配是否具有协同效应,目前尚不清楚。
本研究于2018年3—5月在重庆大学虎溪校区生命科学学院任茂智课题组613真菌实验室进行,目的在于观察雷帕霉素对马铃薯早疫病菌的室内抑制效果,使用透射电镜及扫描电镜在超微结构上揭示雷帕霉素抑菌的机制,并联合甲霜·噁霉灵探索协同抑制效应,以期为安全有效防治马铃薯早疫病提供参考。
1 材料与方法
1.1 菌株及药剂
马铃薯早疫病菌NDL及NBF-5小种由北京北纳创联生物技术研究院提供,该菌株保存在本实验室。将早疫病菌NDL及NBF-5小种接种于PDA培养基上,于25 ℃黑暗条件下培养。
雷帕霉素,美国Selleck Chemicals公司生产;甲霜·噁霉灵,飞翔农资有限公司;马铃薯葡萄糖琼脂培养基(Potato Dextrose Agar,PDA),北京索莱宝公司。
1.2 雷帕霉素处理早疫病菌
雷帕霉素处理早疫病菌的浓度参照用雷帕霉素处理番茄灰霉病菌、油菜菌核病菌等4种植物病原真菌[8]的方法,设置浓度梯度为1、10和50 nmol·L-1。
固体PDA培养基中添加了DMSO(Dimethyl sulfoxide,作为溶剂对照)及前述不同浓度的雷帕霉素。然后,将直径为7 mm的NDL及NBF-5菌块接种于上述PDA培养基上。25 ℃黑暗条件下培养8 d后,测量DMSO对照组及雷帕霉素处理组的菌落直径。数据利用Students t-test和标准差进行统计分析。上述试验每个处理重复3次。
1.3 透射电镜及扫描电镜观察
早疫病菌丝接种于PDA液体培养基中,震荡培养5 d,形成菌丝小球。然后将菌丝小球接种于DMSO对照组及雷帕霉素处理组(20 μmol·L-1)的PDA液体培养基中,25 ℃黑暗培养5 d。5 d后收集菌丝小球,送至重庆医科大学电镜室制片并观察。
1.4 农药甲霜·噁霉灵处理早疫病菌
甲霜·噁霉灵处理早疫病菌的体积浓度参照其使用说明书,设置浓度梯度为0.05 mL·L-1、0.5 mL·L-1和1 mL·L-1。
固体PDA培养基中添加了DMSO(溶剂对照)及前述不同浓度的甲霜·噁霉灵。然后,将直径为7 mm的NDL及NBF-5菌块接种于上述PDA培养基上。25 ℃黑暗条件下培养8 d后,测量DMSO对照组及甲霜·噁霉灵处理组的菌落直径。数据利用Students t-test和标准差进行统计分析。上述试验每个处理重复3次。
1.5 雷帕霉素+甲霜·噁霉灵组合用药处理早疫病菌
将雷帕霉素和甲霜·噁霉灵按不同浓度分别两两组合来处理早疫病菌,得到9种组合。组合用药时,“雷帕霉素(nmol·L-1)+甲霜·噁霉灵(mL·L-1)”的浓度分别为:“1+0.05”“1+0.5”“1+1”“10+0.05”“10+0.5”“10+1”“50 +0.05”“50+0.5”和“50+1”。将两种菌株NDL及NBF-5接种于上述浓度组合固体培养基中,黑暗培养8 d后观察记载菌落直径。采用CompuSyn软件计算CI(Combination index)值[9],药物相互作用关系分别如下:CI<1(协同作用)、CI=1(累加作用)及CI>1(拮抗作用)。
2 结果与分析
2.1 雷帕霉素对早疫病菌的抑制效应
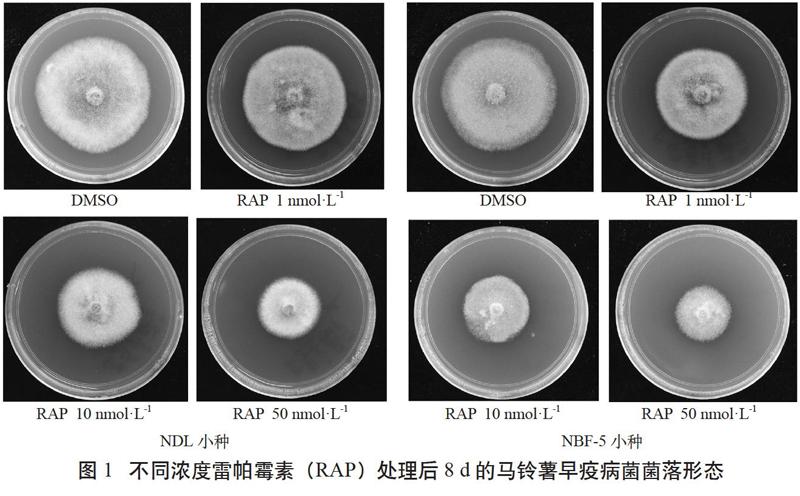
由表1可见,当用雷帕霉素处理早疫病菌菌株NDL时,菌落生长受到显著抑制且呈剂量依赖效应,在以菌株NBF-5为研究对象时也得到相似的结果。
由图1可见,雷帕霉素抑制NDL的IC50值(50%抑制率的浓度)约为50 nmol·L-1,抑制NBF-5的IC50值约为10 nmol·L-1。
2.2 雷帕霉素对早疫病菌超微结构的影响
为揭示雷帕霉素抑制早疫病菌的作用机制,本研究使用扫描电镜及透射电镜分析雷帕霉素对该病原菌NDL及NBF-5两小种超微结构的影响。
据图2扫描电镜观察结果显示:与对照相比,经过雷帕霉素处理的早疫病菌NDL及NBF-5两小种,菌丝节间膨大,隔间距缩短,这可阻碍菌丝延伸[10]。
据图3透射电镜观察结果显示:1)与空白对照组及DMSO对照组相比,用雷帕霉素处理早疫病菌NDL真菌菌丝后,其细胞产生空腔,细胞呈坏死状态;2)细胞内脂滴减少;3)细胞间隔膜发生断裂。在以NBF-5为研究对象时也获得相似结果,即细胞隔膜破裂,细胞空腔增大、坏死等。脂滴在病原菌的各种生理过程中发挥着重要作用,例如,推动细胞生长及病原菌的侵染等[11-14]。雷帕霉素导致细胞内空腔增多且脂滴减少,这可能会致使早疫病菌生长受到抑制及致病力减弱。并且,细胞隔膜在细胞分裂及增殖中作用显著,雷帕霉素可通过破坏细胞隔膜抑制细胞分裂增殖,起到抑菌作用。扫描电镜与透射电镜观察结果表明,雷帕霉素可通过破坏病原菌的超微结构起到抑菌作用。
2.3 甲霜·噁霉灵对早疫病菌的抑制效应
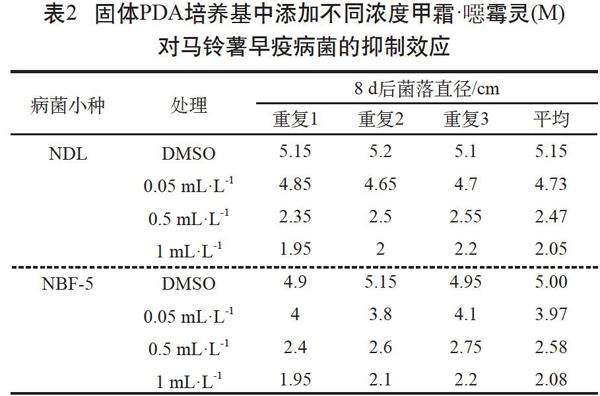
由表2可见,用甲霜·噁霉灵处理早疫病菌株NDL时,随着甲霜·噁霉灵浓度的增加,真菌菌丝生长受到明显抑制。甲霜·噁霉灵抑制NDL和NBF-5的IC50值皆约为0.5 mL·L-1。因此,甲霜·噁霉灵可有效抑制早疫病菌且呈现出剂量依赖效应。
2.4 雷帕霉素+甲霜·噁霉灵组合用药对早疫病菌的协同抑制效应
为探索雷帕霉素与农药联用是否具有协同作用,本研究以甲霜·噁霉灵为农药代表进行相关试验。试验结果发现,当雷帕霉素处理浓度为1~50 nmol·L-1与甲霜·噁霉灵处理浓度为0.05~1.00 mL·L-1组合处理马铃薯早疫病菌株NDL时,双药组合处理NDL的菌落直径明显小于单药处理的菌落直徑,说明双药药效大于单药药效(见表3)。如使用雷帕霉素1 nmol·L-1+甲霜·噁霉灵0.05 mL·L-1组合处理NDL的菌落直径,明显小于雷帕霉素1 nmol·L-1(见表1)或甲霜·噁霉灵0.05 mL·L-1单独处理时的菌落直径(见表2)。其中,雷帕霉素抑菌的IC50值是50 nmol·L-1,甲霜·噁霉灵的IC50值是0.5 mL·L-1,两种药物组合后的IC50值是50 nmol·L-1+0.05 mL·L-1,即双药组合降低了甲霜·噁霉灵使用量9倍。进一步,根据CompuSyn软件计算得到CI<1,说明这两种药物之间存在协同效应。在以早疫病NBF-5为研究对象时也得出相似的结论。
以上试验结果说明,与单药相比,双药联用增强了药效且降低了药物用量。
3 小结与讨论
雷帕霉素为TOR蛋白激酶的第一代抑制剂,雷帕霉素与FKBP12形成的复合体作用于TOR蛋白激酶的FRB结构域[7],从而抑制其活性。已有研究发现,雷帕霉素可导致小麦赤霉病菌菌丝膨大扭曲,细胞超微结构发生变化,对小麦赤病菌具有明显抗菌活性[15]。本研究以马铃薯早疫病菌的两个小种为研究对象,探索了雷帕霉素对早疫病的抑菌效果,发现雷帕霉素能够有效抑制早疫病菌生长;通过透射电镜及扫描电镜揭示雷帕霉素可导致菌丝节间膨大,胞内空腔增多及隔膜破裂等,这些超微结构的明显改变都可导致菌丝生长受阻,达到抑菌效果。
协同作用又称增效作用,是指两种或两种以上药物联合使用时,药效增强。在临床医学中,药物协同作用的原理被广泛应用,如:Aplidin与阿糖胞苷可联合协同治疗白血病[16],大黄素与姜黄素联用协同抑制肝癌细胞生长[17]。同时,一些复合型农药制剂也应用了药物协同作用的原理,如:联合使用氟吡菌胺和霜霉威盐酸盐组成的银法利可有效防治致病疫霉[18]。两种抗真菌药物联合使用,从而产生协同抑菌作用,有效减少药量且大幅度增强药效,是一种简单易行的防治病害的策略。本研究发现,当雷帕霉素与化学农药甲霜·噁霉灵联用时,可有效增强对马铃薯早疫病菌的抑制作用且降低药物使用量,即具有明显的协同效应。
参考文献:
[1] 徐建飞,金黎平.马铃薯遗传育种研究:现状与展望[J].中国农业科学,2017,50(6):990-1015.
[2] 周岱超.马铃薯早疫病季节流行动态及病原菌侵染关键天气条件[D].保定:河北农业大学,2014.
[3] Harrison M D, Venette J R. Chemical control of potato early blight and its effect on potato yield[J]. American Potato Journal, 1970, 47(3): 81-86.
[4] 李雅南,Shah S R A,孙少慧,等.黑龙江省马铃薯叶斑类病害Alternaria属病原组成和分布[J].分子植物育种,2017(3):303-309.
[5] 郑寰宇.马铃薯早疫病菌生物学特性及致病力分化的研究[D].大庆:黑龙江八一农垦大学,2010.
[6] Harrison D E, Strong R, Sharp Z D, et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice[J]. Nature, 2009, 460(7253): 350-356.
[7] Dolinski K, Scholz, C, Muir, R S, et al. Functions of FKBP12 and Mitochondrial Cyclophilin Active Site Residues In Vitro and In Vivo in Saccharomyces cerevisiae[J]. Molecular Biology of the Cell, 1997, 8(11): 2267-2280.
[8] 王彦,徐曦,张立钦,等.雷帕霉素对4种植物病原真菌的抑菌活性[J].农药学学报,2016,18(3):393-396.
[9] Chou T C, Talalay P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors[J]. Advances in Enzyme Regulation, 1984, 22: 27-55.
[10] Zhao C, Waalwijk C, De Wit PJGM, et al. EBR1, a novel Zn 2 Cys 6 transcription factor, effects virulence and apical dominance of the hyphal tip in Fusarium graminearum[J]. Mol Plant-Microbe Interact, 2011, 24(12): 1407-1418.
[11] Bago B, Zipfel W, Williams R M, et al. Translocation and utilization of fungal storage lipid in the arbuscular mycorrhizal symbiosis[J]. Plant Physiol, 2002, 128(1): 108-124.
[12] Kobae Y, Gutjahr C, Paszkowski U, et al. Lipid droplets of arbuscular mycorrhizal fungi emerge in concert with arbuscule collapse[J]. Plant Cell Physiol, 2014, 55(11): 1945-1953.
[13] Radulovic M, Knittelfeldero, Cristobal-Sarramian A, et al. The emergence of lipid droplets in yeast: current status and experimental approaches[J]. Curr Genet, 2013, 59(4): 231-242.
[14] Stehr M, Elamin A A, Singh M. Cytosolic lipid inclusions formed during infection by viral and bacterial pathogens[J]. Microb Infect, 2012, 14(13): 1227-1237.
[15] Yu F, Gu Q, Yun Y, et al. The TOR signaling pathway regulates vegetative development and virulence in Fusarium graminearum[J]. New Phytologist, 2014, 203(1): 219-232.
[16] Jia J, Zhu F, Ma X, et al. Mechanisms of drug combinations: interaction and network perspectives[J]. Nat Rev Drug Discov, 2009, 8(2): 111-128.
[17] 雷湘,李娟,陳科力.大黄素与姜黄素联用对肝癌细胞BEL-7402生长抑制的协同作用[J].中国医院药学杂志,2009,29(12):971-973.
[18] Samoucha Y, Hugelshofer U, Gisi U. Effects of disease intensity and application type on efficacy and synergy of fungicide mixtures against Phytophthora infestans[J]. J Phytopathol, 2010, 120(1): 44-52.
(责任编辑:丁志祥)
收稿日期:2019-12-29
基金项目:政府间国际科技创新合作重点专项(National key R&D program of China)(2017YFE0115500);重庆市基础研究与前沿探索专项(cstc2018jcyjAX0753);国家自然科学基金项目(31972469);成都农业科技中心地方财政专项(NASC2019TI13);中央科研单位基本科研业务费项目(19-001-09);重庆市科技攻关项目(cstc2015SHMS-ZTZX80003);中央高校基本科研业务费研究生科研创新项目(2018CDYJSY0055)。
作者简介:吕秀兰(1993—),女,湖北恩施人,重庆大学生命科学学院生物学专业2017级在读硕士研究生,研究方向为植物病理学。E-mail: 1392814143@qq.com。※为通信作者,E-mail: renmaozhi01@caas.cn。
