黄土高原刺槐人工林根际和非根际土壤磷酸酶活性对模拟降水变化的响应
2020-04-27时新荣1袁志友1
刘 迪, 邓 强, 时新荣1,, 袁志友1,
(1.西北农林科技大学, 黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌 712100; 2.西北农林科技大学 林学院, 陕西 杨凌 712100;3.中国科学院 水利部 水土保持研究所, 黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨凌 712100)
人类活动导致的全球变暖使降水格局发生改变,温度上升增加了海洋和陆地的蒸腾,导致降雨增加,同时也会增加蒸散量,其结果是进一步减少土壤的湿润度,干旱地区温度升高可能会使本地区更加干旱[1]。多个大气循环模型预测未来极端降水事件和极端干旱事件发生的频率会显著增加[2]。降水变化影响土壤物理化学性质[3]、分解者活动和有机物质分解速率,从而改变土壤养分动力学和生物有效性[4-5]。在全球尺度上,温度上升导致连续无雨日数和连续降雨日数等极端降水指数有显著升高的趋势;在区域尺度上,尤其是生态环境脆弱的地区,极端降水事件更加受到广泛关注[6]。
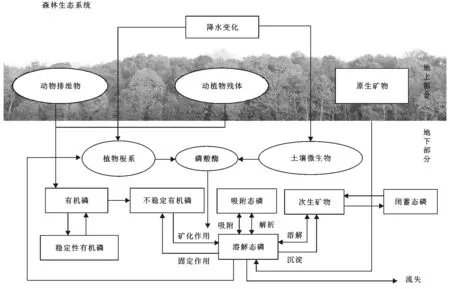
图1 降水变化影响土壤有机磷的概念
土壤磷酸酶(Soil phosphatase)是催化土壤有机磷加快矿化的一类水解酶的合称,主要来自植物根系和土壤微生物,也有少部分来自动植物残体[7]。土壤磷酸酶活性是衡量土壤有机磷矿化速率和磷素有效性的重要指标[8],有学者认为,在磷酸酶缺乏的情况下,土壤有机磷的释放需要几百年的时间[9]。根据已有研究,土壤有机磷主要来自动植物残体和动物排泄物,其在土壤中可进行不断转化,降水对土壤磷酸酶以及有机磷矿化过程影响机制如图1所示,磷酸酶的催化作用在此过程中必不可少[10],并且与植物根系相互作用,而细根(直径≤2 mm)因大的根系表面积和强的活性,与磷酸酶活性关系密切[11],研究发现,细根生物量越大,根系分泌速率越高[12]。磷酸酶活性极易受环境因子变化的影响,其中与降水变化的关系仍不明确,其对降水变化的响应因生态环境和植被类型的不同而具有明显的差异性[7-13]。磷酸酶活性与土壤有机磷的关系复杂,有机磷作为磷酸酶的底物,其含量升高有利于磷酸酶的产生[14],而过多的磷酸酶又会反作用于有机磷[15]。目前,有关不同降水变化对黄土高原森林生态系统土壤磷酸酶活性影响的野外试验还未见报导[16],其余地区已有的相关试验还未考虑在多个降水处理强度下,尤其是极端干旱和极端降水条件下土壤磷酸酶的响应规律[17]。由于土壤中的磷与水分密切相关,可以预测,在干旱的黄土高原区的土壤磷酸酶活性对降水变化的响应会与其他生态系统有所不同。
黄土高原由于其脆弱的生态环境,极容易受到未来降水格局变化的影响,降水已经成为黄土高原半干旱区的主要限制因素,据近年来的统计结果显示,黄土高原大部分区域极端干旱发生频率有升高的趋势[18]。刺槐(RobiniapseudoacaciaLinn.)具有较强的抗旱能力,是黄土高原主要造林树种之一,在该地区分布非常广泛,但近年来因刺槐林立地条件不适宜而逐渐出现树种结构单一、生态效益低下、林分退化等问题[19]。在全球降水变化背景下研究刺槐林地土壤磷酸酶的变化方向对黄土高原地区植被恢复和水土保持工作有着重要指导意义。本研究以黄土高原35 a生刺槐人工林为研究对象,进行野外控制穿透雨试验,旨在研究:(1) 根际和非根际土磷酸酶活性对7个穿透雨变化强度的响应规律及原因;(2) 比较根际土与非根际土间磷酸酶活性的差异;(3) 根际和非根际磷酸酶活性在土层间的变化特征;(4) 探索影响根际和非根际土磷酸酶活性的主要因子及影响特征。为研究全球气候变化下森林土壤有机磷矿化潜力、磷素生物转化方向及其强度提供依据。
1 材料与方法
1.1 研究区概况
试验地点位于陕西省咸阳市永寿县槐平林场(34°83′N,108°09′E),海拔1 382 m,属暖温带半湿润大陆性季风气候,冬季漫长干寒,夏季短暂温湿,年平均气温10.8℃,年平均降雨量602 mm,试验期间月平均降雨量和月平均气温如图2所示。土壤母质为黄土,土壤类型为黄绵土、褐土和黑垆土。土壤基本理化性质如下:总有机碳19.1 g/kg,总氮1.10 g/kg,总磷0.67 g/kg,速效磷16.1 mg/kg,速效钾0.14 g/kg,土壤含水量12.0%,pH值8.07。槐平林场在植被区划分类型上属暖温带落叶阔叶林区域,乔木层结构较单一,现主要为林龄在30~35 a的刺槐人工林,其林下灌木主要有茅梅(RubusparvifoliusLinn.)、南蛇藤(CelastrusorbiculatusThunb.)、山梅花(PhiladelphusincanusKoehne)等;草本层主要包括三脉紫菀(AsterageratoidesTurcz.)、拉拉藤(Humulusscandens(Lour.)Merr.)、老鹳草(Geraniumwilfordiimaxim.)、天名精(CarpesiumabrotanoidesL.)、商陆(PhytolaccaacinosaRoxb.)等。

图2 试验期间研究区月降水量和气温变化
1.2 样地设置
我们选择黄土高原刺槐人工林,于2015年6月在该研究样地进行穿透雨控制试验,共设置21块15 m×15 m的样方,样方间隔不小于100 m,分成7组处理模式,分别为减水80%(拦截穿透雨面积为林地投影面积的80%)、减水60%(拦截穿透雨面积为林地投影面积的60%)、减水20%(拦截穿透雨面积为林地投影面积的20%)、对照、增水20%(施加减水20%样地拦截的穿透雨)、增水60%(施加减水60%样地拦截的穿透雨)、增水80%(施加减水80%样地拦截的穿透雨),每个处理模式设置3个重复样方。减水处理采用透光率90%的PE薄膜作为遮雨膜,减水样方按照减水比率×样方林地投影面积布设相应面积的遮雨膜,根据样方地势走向和坡度,设置PVC水管收集遮雨膜上的雨水,每次降水后,将收集的雨水均匀转移到相同比率的增水样方中,来实现一定比率降水的增加和减少。
1.3 植物样和土样采集
本次研究于2017年7月进行土壤样品采集,在每个处理样地内,按照五点取样法,将50 cm×50 cm样方框内植物绿色部分齐地面剪下,分类装入信封袋并标记,75℃烘干至恒重后称重。用直径9 cm根钻分层采集0—10 cm,20—40 cm,40—60 cm的土样,采用抖落法挑出30 g根际土与30 g非根际土,分别装入干净聚乙烯自封袋,4 ℃冰箱保存,用于测定碱性磷酸酶活性,其余土样去除可见杂物后,取约30 g装入铝盒,测定土壤含水量,约200 g自然风干,测定土壤理化指标。剩余土样用0.2 mm筛子冲洗,拣出细根(直径≤2 mm),75℃烘干至恒重后称重。
1.4 指标测定方法
总有机碳测定采用重铬酸钾外加热法,总氮测定采用凯式定氮法,总磷测定采用酸解—钼锑抗比色法,速效磷采用NaHCO3浸提—钼锑抗比色法测定,速效钾测定采用醋酸铵浸提—火焰原子分光光度法测定,根际与非根际碱性磷酸酶测定采用对硝基苯磷酸盐法,以单位质量土壤样品中碱性磷酸酶在单位时间内产生的酚含量表示,以下磷酸酶均指的是碱性磷酸酶(ALP),用烘干—称重法测定土壤重量含水量,用pH计测定土壤pH值[20],土壤有机磷测定采用Hedley分级法[21]。
1.5 数据处理
采用单因素方差分析(One-way ANOVA)检验7种降水处理间土壤含水量、pH值、有机磷含量、林下优势科植物地上生物量、总地上生物量、细根生物量、根际和非根际磷酸酶活性的差异,采用线性回归分析(Linear regression analysis)评价土壤含水量、pH值、有机磷含量、林下优势科植物地上生物量、总地上生物量、细根生物量、根际和非根际磷酸酶活性的关系以及磷酸酶土层间递减率和土层的关系。
2 结果与分析
2.1 7种模拟降水处理下生物量和土壤理化特征
降水处理影响林下植被地上生物量和细根生物量,根据样地植被调查数据,将刺槐人工林林下植物分为茜草科(RubiaceaeJuss.)、菊科(AsteraceaeBercht. & J.Presl.)、蔷薇科(RosaceaeJuss.)和杂草类4个大类。其中茜草科植物地上生物量在不同处理间差异显著,菊科地上生物量、蔷薇科地上生物量、总地上生物量和细根生物量处理间差异均达到极显著水(p<0.01),而杂草类处理间差异不显著,各科地上生物量、总地上生物量和细根生物量均随降水增加表现出升高的趋势(表1)。
不同降水处理对研究区土壤含水量和有机磷含量有明显影响,0—20 cm土层土壤含水量减水60%,80%和增水80%处理间差异显著,40—60 cm土层土壤含水量在减水60%、增水60%和对照间差异显著;不同处理间0—20 cm土壤有机磷含量差异接近显著水平(p<0.08),而土壤pH值、20—40 cm,40—60 cm土壤有机磷含量差异不显著。减水样地土壤含水量较对照样地共减少7.32%,增水样地土壤含水量较对照样地共增加17.7%。土壤含水量和有机磷含量随降水增加表现出升高的趋势(表2)。

表1 不同降水处理样地林下植物群落组成以及生物量特征

表2 7种降水处理对土壤理化性质的影响
2.2 降水变化对根际土和非根际土磷酸酶活性的影响
研究区7个降水梯度上根际土磷酸酶活性和非根际土磷酸酶活性的变化范围分别是103~513 mg/(kg·h),78.7~443 mg/(kg·h)。增水处理对黄土高原刺槐人工林0—20 cm,20—40 cm土层根际土磷酸酶活性影响明显,在3个增水梯度上随降水增加呈增大趋势,但是在3个减水梯度上,根际土磷酸酶活性没有表现出一致的规律。0—20 cm土层减水60%处理的根际土磷酸酶活性最小,为273 mg/(kg·h),增水80%处理活性达到最大,为513 mg/(kg·h);20—40 cm土层减水20%处理其活性最小,为128 mg/(kg·h),增水80%处理其活性达到最大,为315 mg/(kg·h)。增水处理的土壤磷酸酶活性在根际与非根际土间差异不显著,0—20 cm土层减水处理的土壤磷酸酶活性在根际与非根际土间差异显著,20—40 cm,40—60 cm差异均不显著,7种降水处理间0—20 cm土层根际土磷酸酶活性差异极显著,20—40 cm土层差异显著,40—60 cm土层差异不显著(图3)。随着降水增加,根际土磷酸酶在采样的3个土层的递减率呈现下降的趋势,0—20 cm至20—40 cm土层和20—40 cm至40—60 cm土层的变化范围分别是38.7%~61.0%,25.0%~57.9%(图4)。
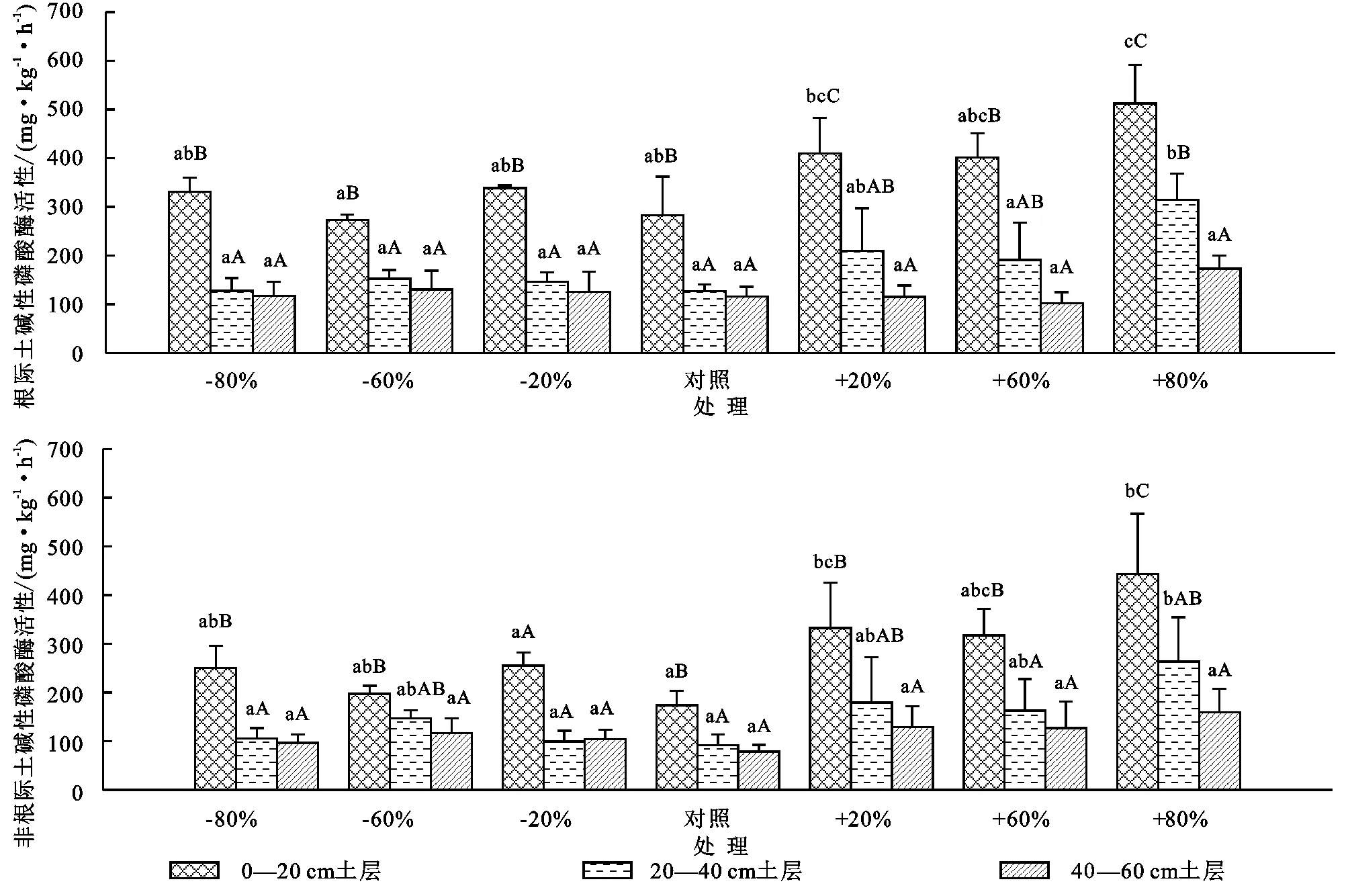
注:不同大写字母表示不同土层间差异显著(p<0.05),不同小写字母表示不同降水处理间差异显著(p<0.05),下同。
黄土高原刺槐人工林0—20 cm,20—40 cm土层非根际土磷酸酶活性随降水增加表现出先减小后增大的趋势,3个梯度上增水处理的非根际土磷酸酶活性显著高于减水处理,0—20 cm土层增水样地(增水20%,60%,80%)比减水样地(减水20%,60%,80%)的非根际磷酸酶活性分别高77.0,120,193 mg/(kg·h);20—40 cm土层增水样地(增水20%,60%,80%)比减水样地(减水20%,60%,80%)分别高80.0,15.5,158 mg/(kg·h)。7种降水处理间0—20 cm土层非根际土磷酸酶活性差异显著,20—40 cm土层接近显著水平,40—60 cm土层差异不显著(图3)。与根际土磷酸酶土层间递减率不同,非根际土磷酸酶在3个土层间的递减率随着降水增加总体呈现出上升趋势,0—20 cm至20—40 cm土层和20—40 cm至40—60 cm土层的变化范围分别是8.50%~44.9%,2.07%~37.3%(图4)。
2.3 不同土层磷酸酶活性及土壤理化特征
研究区域20—40 cm土壤含水量和有机磷含量分别比0—20 cm土壤低25.3%和41.1%,pH值高3.66%;40—60 cm土壤含水量和有机磷含量分别比20—40 cm土壤低14.3%和9.92%,pH值高0.26%。在土壤纵向分布上,土壤含水量和有机磷含量随土层深度增加均呈下降趋势,其中土壤含水量土层间差异达到极显著水平(p<0.01)。pH值则呈上升趋势,但土层间差异没有达到显著水平。
不同土层根际土磷酸酶活性和非根际土磷酸酶活性差异均显著。随土层深度增加,根际土和非根际土磷酸酶活性表现出明显的下降趋势。研究区域7个降水处理下0—20 cm,20—40 cm,40—60 cm土壤根际土磷酸酶活性平均分别为364 mg/(kg·h),181 mg/(kg·h),165 mg/(kg·h);0—20 cm,20—40 cm,40—60 cm土壤非根际土磷酸酶活性平均分别为281 mg/(kg·h),149 mg/(kg·h),115 mg/(kg·h)。20—40 cm土壤根际土磷酸酶活性和非根际土磷酸酶活性分别比0—20 cm土壤低50.2%,46.7%;40—60 cm土壤根际土磷酸酶活性和非根际土磷酸酶活性分别比20—40 cm土壤低30.7%,22.7%(图3)。根际土和非根际土磷酸酶在3个土层间的递减率随土层深度增加而显著减小,分别比深20 cm处土层的递减率高3.36%和6.87%。根际土磷酸酶土层递减率要高于非根际土,采样的3层土中,0—20 cm至20—40 cm土层的根际土磷酸酶递减率比非根际土高26.1%,第20—40 cm至40—60 cm土层则高31.7%(图4)。

图4 7种降水处理对不同土层土壤磷酸酶递减率的影响

图5 碱性磷酸酶活性与土壤有机磷含量、土壤含水量的关系
2.4 土壤磷酸酶活性与土壤理化性质、生物量的相关性
研究区根际土磷酸酶活性与土壤含水量呈极显著正相关(图5),与土壤有机磷呈显著正相关。非根际土磷酸酶活性与土壤含水量呈正相关关系(p=0.011),与土壤有机磷呈显著正相关。
根际土磷酸酶和非根际土磷酸酶活性与pH值呈显著负相关(p<0.01)。根际土磷酸酶活性与非根际土磷酸酶活性呈极显著正相关(p<0.001),非根际土磷酸酶随根际土磷酸酶升高而升高(图6)。
就不同科植物来说,根际土磷酸酶活性与蔷薇科植物地上生物量呈极显著正相关,与地上生物量相关性接近显著水平(p=0.06)。非根际土磷酸酶活性与蔷薇科植物呈极显著正相关关系,与细根生物量显著正相关,与总地上生物量相关性接近显著水平(p=0.065)。研究区磷酸酶活性与茜草科、菊科和杂草类均无显著相关关系(表3)。

图6 根际土碱性磷酸酶活性与非根际土碱性磷酸酶活性的关系

表3 生物量与磷酸酶活性的相关性分析
3 讨论与结论
(1)根际和非根际土壤磷酸酶活性对降水变化的响应。研究区根际土磷酸酶活性和非根际土磷酸酶活性的变化范围均在黄土高原已有研究结果的变化范围之内[22],但低于中国东北、华南林地以及地中海灌丛地[14,23-24]。这可能是因为黄土高原气候干燥,降水季节分配不均,加之黄土土质疏松,使该地区极易发生水土流失,而土壤水分和有机质是土壤酶活性的重要影响因子[25],干旱的气候条件和较贫瘠的土壤条件可能是该区磷酸酶活性低于前文提到的部分地区的原因。
根际土和非根际土磷酸酶活性在增水梯度上均随降水增加表现出升高趋势,这与已有盆栽试验得出的磷酸酶活性随降水增加先升高后下降的结果不一致[26],原因可能是本试验属于野外定位试验,野外环境因素较复杂,与室内试验结果往往不一致,黄土高原属于半干旱地区,短期增水没有使土壤达到过湿的效果。降水一方面增加了土壤微生物和植物根系活力,刺槐林植物根系、菌根和植物残留物能够渗出某些不稳定化合物,这些物质和土壤湿度的增加可以刺激微生物产生酶[27],另一方面提高土壤有机物质含量,磷酸酶可作用底物增加[26],这可能是穿透雨增加梯度上磷酸酶活性逐渐升高的原因。
在研究区设置的3个降水减少梯度上,减水20%处理磷酸酶活性要高于对照处理,这可能是因为遮雨20%的轻度干旱环境既保持了一定土壤水分,又增加了土壤的透气性,为植物根系和微生物活动提供了一个短期的良好环境[17],减水60%处理下磷酸酶活性显著降低,这与以往研究结果相同[28]。而极端干旱的减水80%处理下土壤磷酸酶活性有少量增加,原因尚不明确,但是明显低于增水处理,这可能是因为植物和共生微生物缓解短期干旱胁迫的一种适应机制,如丛枝菌根真菌具有磷酸酶活性,能增加寄主对磷的利用,增强其抗旱性[29],而丛枝菌根真菌对寄主具有选择性,蔷薇科植物和豆科植物根系极易被丛枝菌根真菌侵染,试验结果表明蔷薇科植物地上生物量与磷酸酶活性显著相关;隔离降雨后土壤水分减少,使土壤溶液中的可溶性磷素减少,而低磷环境又可以刺激根系和外生菌根分泌磷酸酶来改善植物的养分状况[30]。非根际土磷酸酶活性在干旱梯度上随降水增加而下降,可能与稀释作用有关,但不管是根际土还是非根际土磷酸酶,其在干旱处理下的活性明显低于增水处理,干旱土壤的土壤含水量和有机质含量较低,磷酸酶可作用底物减少,土壤微生物活性下降,元素周转速率下降,植物代谢缓慢,使干旱环境中土壤磷酸酶显著低于湿润环境[31]。
(2)根际土与非根际土间磷酸酶活性的比较。不同降水处理对非根际土磷酸酶活性的影响与根际土相同,非根际土酶活性受到根际土酶活性的强烈影响,与根际土磷酸酶活性呈明显的正相关关系,并且活性低于根际土。土壤酶主要来自土壤微生物、植物根系和动植物残体,在土壤环境中,植物与其根系周围土壤的物质交换更频繁,植物自身代谢以及与植物根系共生的微生物分泌的胞外酶首先进入根际土壤,使得根际土中酶活性比非根际更高[15]。本研究中不同降水处理对0—20 cm土层的根际土磷酸酶活性影响最显著,对20—40 cm土层影响较低,对再深土层则没有影响,这与Sardans等[32]在地中海圣栎林的穿透雨试验中得出的降水减少对0—15 cm土层土壤磷酸酶活性影响更明显、在空间分布上随土层深度增加影响逐渐减小的结论一致,但与王楠楠等[7]得出的降水变化对土壤酶活性没有显著影响的结果不一致,这可能是因为在黄土高原刺槐人工林这种特定环境下,1~2 a的降水处理虽然能够影响土壤磷酸酶活性,但还仅能影响0—40 cm土层的土壤酶活性,对更深土层的土壤酶活性及其与之有关的各项理化指标(如土壤含水量、微生物活性和土壤有机质含量)影响还非常小。
磷酸酶土层间递减率在根际土和非根际土间有不一样的规律,根际土磷酸酶递减率随降水增加而下降,这有可能是水分增大了磷酸酶的扩散速率,减小了土层间的差异,而非根际土磷酸酶递减率则随降水增加而上升,说明水分加速了土壤磷酸酶从根际到非根际的土层内的迁移速度,但是土层间的迁移速度变化不大。
(3)降水影响不同土层根际和非根际土壤磷酸酶活性的因素。黄土高原刺槐人工林根际与非根际土磷酸酶活性随土层深度增加而显著减小,磷酸酶土层递减率也表现出显著减小的趋势,这与以往研究结果一致[31-32]。pH值呈增大趋势,但是土层间差异不显著。磷酸酶活性在空间上与土壤自身的异质性有相当大的关系,本研究中土壤含水量和有机磷随土层深度增加有明显的下降趋势,水分利用性直接影响土壤微生物活性和土壤有机质含量,研究表明,陆地生态系统中的土壤资源和植被覆盖是异质的,土壤水分、微生物活性、酶活性和养分含量都会受到异质性的影响[33]。
土壤酶活性受到多种因子的共同作用,各土层土壤碱性磷酸酶并没有随pH值升高而升高,而是与土壤含水量和土壤有机磷呈显著的正相关关系。土壤中磷酸酶活性受土壤湿度极大影响,Brockett等人认为,土壤含水量是影响土壤酶活性最主要的因子,且酶活性随着土壤湿度的增大而增强[34]。土壤磷酸酶与有机磷的关系尚不明确,一部分观点认为酶具有专一性,有机磷是磷酸酶的水解底物,有机磷含量的增加会刺激磷酸酶的产生[14],另一观点认为磷酸酶加快了有机磷的矿化速率,同时高的无机磷含量会对磷酸酶产生负反馈作用,两者呈负相关关系[15-35],本研究结果与前者一致,这可能是因为在黄土高原的短期降水虽然增加了有机磷含量,但仍未达到室内试验中人为添加的过量水平。磷酸酶活性随细根生物量的增加而增加,与王靓等[11]的研究结果一致。研究表明,细根生物量越大根系分泌速率越高[12]。地上生物量与磷酸酶呈正相关关系,这可能是因为植物需要一定的磷酸酶来帮助吸收土壤中的磷,生物量增加,物质循环进程加快,有利于根系生长和微生物活动产生更多磷酸酶,但生态系统物质循环过程是相当复杂的,还需更具体的研究来解释生物量与磷酸酶的关系。磷酸酶与蔷薇科植物地上生物量呈极显著的正相关关系,与茜草科植物相比,蔷薇科植物根系更容易与菌根真菌共生形成菌根,有关内蒙古草原的调查研究发现,在所有被调查的植物中,蔷薇科植物的丛枝菌根侵染率最高[36],其不仅能增加植物的抗旱能力,还能分泌磷酸酶促进植物对磷素的吸收。
综上所述,与短期降水减少相比,短期降水增加对黄土高原土壤磷酸酶活性的影响更显著,且具有积极作用,这种作用随土层深度增加而减小,影响深度约至40 cm。降水变化通过影响土壤含水量、土壤有机磷和地上植物改变土壤磷酸酶活性,进而对有机磷矿化产生影响。研究降水变化对土壤磷酸酶的影响对黄土高原土壤磷循环研究和植被恢复具有重要的理论意义和应用价值。
