水稻和两种野生植物对酸性硫酸盐土耐性及矿质元素吸收*
2020-04-25张玲玉赵学强李家美沈仁芳
张玲玉,赵学强†,李家美,沈仁芳
水稻和两种野生植物对酸性硫酸盐土耐性及矿质元素吸收*
张玲玉1,3,赵学强1,3†,李家美2,沈仁芳1,3
(1. 土壤与农业可持续发展国家重点实验室(中国科学院南京土壤研究所),南京 210008;2. 河南农业大学生命科学院,郑州 450002;3. 中国科学院大学,北京 100049)
酸性硫酸盐土是发育于还原性硫化物成土母质的一种恶劣土壤类型,虽然其分布区气候条件十分有利于水稻生长,但由于土壤条件限制,水稻生产潜力难以发挥。为筛选出适应该土壤的水稻品种,在酸性硫酸盐土(pH 3.69)上种植了240个水稻品种,但是所有水稻品种均未获得经济产量,却发现两种野生植物在该土壤中生长茂盛。经形态和分子鉴定,两种野生植物为野荸荠()和五棱飘拂草()。进一步分析了水稻和两种野生植物的根际和非根际土壤性质、根部和地上部矿质元素含量。结果表明,根际土壤pH、交换性铝和有效锰含量不能解释水稻和两种野生植物耐性差异,锰毒和缺氮亦不是酸性硫酸盐土限制植物生长的关键因子,而植物地上部矿质元素含量与植物耐性差异相关。铝毒、铁毒和较低的养分(磷、钾、钙、镁)吸收是酸性硫酸盐土中水稻生长不良的主要因子。相反,两种野生植物对铝毒和铁毒的耐性较强,对养分吸收能力较强,所以生长良好。
酸性硫酸盐土;水稻;野荸荠;五棱飘拂草;金属毒害;养分缺乏
酸性硫酸盐土是指发育于富含还原性硫化物成土母质的土壤,一般土壤沉积层中含有硫化铁氧化或未被氧化的致酸层和潜在致酸层,是世界范围内最恶劣的土壤类型之一[1-2]。全球范围内,酸性硫酸盐土超过了1 700万hm2,其中650万hm2分布于亚洲,450万hm2在非洲,300万hm2在澳洲,300万hm2在拉丁美洲,23.5万hm2在欧洲(主要是芬兰和瑞典),10万hm2在北美洲,主要分布于热带亚热带滨海地区[3-4]。我国酸性硫酸盐土面积约10.6万hm2,主要分布于广东、广西、海南、福建、台湾等省(区)的入海河口及海岸沿线,其中广东省分布面积最大,约占全国酸性硫酸盐土总面积的60%左右[4-5]。我国是农用耕地资源严重缺乏的国家,耕地面积还在不断缩减,粮食安全问题日趋严重[6]。酸性硫酸盐土分布地区水热资源丰富,适宜大多数作物生长。科学利用已被开垦的酸性硫酸盐土类型农田,维持被开垦酸性硫酸盐土地区农业可持续发展,对于粮食安全和生态环境保护均具有重要意义。
酸性硫酸盐土被开垦或处于氧化环境时,其中的硫铁矿物质与空气接触发生氧化,释放大量硫酸,形成强烈酸害,严重影响区域内动植物生长和人类健康[7]。广东省地处我国大陆最南端,属东亚季风区,是中国光、热和水资源最丰富的地区之一,气候条件非常适合水稻种植。目前,位于珠江三角洲的大部分酸性硫酸盐土已被开垦为稻田。被围垦的酸性硫酸盐土稻田,由于地块间土壤环境状况不同,水稻生长参差不齐,严重者在插播数小时内秧苗全部枯萎,一天后死亡;危害较轻者植株矮小,生育后期全株发黑,不实率很高[8]。当农民收益较低时,被开垦的酸性硫酸盐土农田易被弃用,弃用的酸性硫酸盐土农田会对周围环境产生恶劣影响。由于土壤条件的限制,酸性硫酸盐土上的水稻生产潜力得不到发挥[9]。增强水稻对土壤逆境胁迫的抗性是发挥酸性硫酸盐土上水稻生产潜力的关键。
在以往酸性硫酸盐土研究中,前人对酸性硫酸盐土的成土条件、矿物转化、土壤化学特征、水分影响机制、作物生长限制因子及改良措施进行了诸多研究[9-13]。然而,很少有研究者关注酸性硫酸盐土围垦农田中水稻与野生植物的耐逆性和养分吸收差异。在酸性硫酸盐土上,虽然水稻生长不良,甚至死亡,但是有的野生植物却能适应该环境,生长非常茂盛[14]。硫酸根含量过高、铝毒、锰毒、酸害、缺磷、缺钾被认为是广东省酸性硫酸盐土水稻产量的主要限制因子[9]。耐逆性和对矿质元素吸收的差异可能是水稻和野生植物对酸性硫酸盐土适应能力差异的原因,但是具体哪种因素占主导,仍不清楚。
为了探讨水稻与野生植物适应酸性硫酸盐土能力和矿质元素吸收之间的关系,在同一块酸性硫酸盐土稻田中,采集了水稻和两种生长茂盛的野生植物样品及其非根际、根际土壤样品,分析了植物根和地上部矿质元素含量及土壤化学性质。本研究试图从植物矿质元素吸收角度解释水稻与两种野生植物适应酸性硫酸盐土能力差异的原因,以便能合理地重点补充酸性硫酸盐土上水稻生长需要的养分,或者为培育新的耐性水稻品种提供信息。
1 材料与方法
1.1 研究区概况
采样地为台山市冲蒌镇八家村(22°06′55.29″N,112°46′32.10″E),属于广东省江门市,位于珠江三角洲西南部。该地区酸性硫酸盐土稻田酸度跨度大,各地块水稻产量差异亦较大。本研究选取了一种酸度较大的地块。根据调研发现,由于该地块种植的水稻长势差、产量低,尤其在降雨少的年份,甚至颗粒无收,因此当地农户将此地块闲置。
1.2 试验设计
2018年在该地块种植了240个水稻()品种。4月19日在当地正常无酸害田块进行水稻育秧,5月17日在酸性硫酸盐土上插秧,试验区总面积约120 m2,土壤性质均一,每个水稻品种种植一行(100 cm × 40 cm),每行6穴,每穴2株,每个水稻品种共12株。整个试验区分为5列,每列48个水稻品种,列与列之间留0.5 m宽度,便于田间调查和管理。2018年8月23日调查采样。根据当地农民施肥习惯,每公顷施用750 kg复合肥(N-P2O5-K2O=18-12-10,氮磷钾总含量大于等于400 g·kg–1),全部以基肥随翻地施入。试验目的是筛选耐酸性硫酸盐土的水稻品种。在水稻生长后期,水稻长势较差,有的品种甚至枯萎,最终所有水稻品种均未获得经济产量。在保护行和两行水稻中间位置,观察到两种野生植物生长非常茂盛,无任何毒害症状。
2018年4月3日,种植水稻前,采集了试验田块(未淹水)的基础土样,测定的土壤基础理化性状如下:pH 3.69,碱解氮175 mg·kg–1,有效磷9.56 mg·kg–1,速效钾102 mg·kg–1。2018年8月23日,在试验区随机选择了三个距离较远的位置,在每个地点采集了3株长势较差的水稻根、地上部、根际土壤和非根际土壤样品,每个地点的样品混合后作为一个重复,共3个重复,每个重复是3株水稻的混合样。同时,采集两种野生植物的根、地上部、根际、非根际土壤样品。由于两种野生植物混杂地生长在一起,根系交错,两种野生植物根际土壤无法区分,最后选择将两种野生植物根际土壤视为一种类型根际土壤,后面称之为野生植物根际土壤,并采集了对应的非根际土壤。植物样品取回实验室后,用蒸馏水冲洗干净,将地上部与根分离,105℃杀青30 min,80℃烘干至恒重。随后,用不锈钢粉碎机将烘干样品粉碎,用于测定植物体内矿质元素含量。同时,根际土壤与非根际土壤分别风干、研磨,过10目和100目的筛子,用于测定土壤化学性质。
1.3 分析项目与方法
在分析各种指标时,每个样品测定2个平行。土壤pH用电位法,水土比2.5︰1测定(PB-21,Sartorius,德国);土壤碱解氮采用碱解扩散法测定;土壤有效磷采用HCl-NH4F浸提—钼锑抗比色法测定;速效钾采用醋酸铵浸提—火焰光度计测定;植物磷(P)、钾(K)、钙(Ca)、镁(Mg)、硫(S)、铁(Fe)、锰(Mn)、铝(Al)、钠(Na)采用硝酸-高氯酸(4︰1)消煮,电感耦合等离子体发射光谱仪(ICP-AES,Optima 8000,PerkinElmer,美国)测定;植物全氮采用硫酸-过氧化氢消煮,靛酚蓝比色法测定;土壤交换性铝采用1 mol·L–1KCl浸提,有效锰采用1 mol·L–1NH4OAC浸提,电感耦合等离子发射光谱仪(ICP-AES,Optima 8000,PerkinElmer,美国)测定;植物体pH采用食品有效酸度方法测定[15]。
野生植物鉴定先请植物分类学家对两种野生植物进行形态学鉴定,同时进行分子鉴定。采用TreliefTMPlant Genomic DNA Kit试剂盒(通用型,北京擎科新业生物技术有限公司),提取了两种野生植物的DNA,进行PCR,分别扩增ITS(内转录间隔区,Internal Transcribed Spacer)序列和ITS2序列,引物和PCR条件参考文献报道[16]。其中ITS序列扩增引物为5afwd:5′-CCTTATCATTTAGAGGAAGG AG-3′;4rev:5′-TCCTCCGCTTATTGATATGC-3′。ITS序列PCR扩增程序:94℃ 5 min;94℃ 1 min,50℃ 1 min,72℃ 1.5 min + 3 s/cycle,30个循环;72℃ 7 min[16]。ITS2序列扩增引物为S2F:5′-ATGC GATACTTGGTGTGAAT-3′;S3R:5′-GACGCTTCTC CAGACTACAAT-3′。ITS2序列PCR扩增程序:94℃5 min;94℃30 s,56℃30 s,72℃ 45 s,40个循环;72℃ 10 min[16]。PCR产物送金斯瑞生物科技有限公司进行测序,获得的核酸序列在美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI)核酸数据库,用基于局部序列比对算法搜索工具(Basic local alignment search tool,BLAST)进行序列比对,将与PCR产物序列相似度高的核酸序列下载,采用MEGA7.0软件中的CLUSTAL W程序对多条序列进行多重比对,最大似然法(Maximum likelihood,ML)构建分子系统进化树,用来分子鉴定两种野生植物。
1.4 数据处理
试验数据为3个重复的平均值,每个重复为2个测试平行的平均值。采用WPS2019(Version 11.1.0)整理和做图,差异显著性分析采用SPSS19.0统计软件中单因素方差分析(ANOVA)方法。
2 结 果
2.1 两种野生植物的鉴定
分别将两种植物的ITS和ITS2的核酸序列提交到NCBI(https://www.ncbi.nlm.nih.gov/)核酸数据库上,分别获得序号MK295082(BQ1,ITS)、MK295083(PFC1,ITS)、MK295084(PFC1,ITS2)、MK295085(BQ1,ITS2),建立了分子系统进化树(图1)。系统进化树中分别用BQ1和PFC1代表发现的两种野生植物,括号中编号为NCBI核酸数据库的序列编号。无论是用ITS还是ITS2构建分子进化树,BQ1均与距离最近(图1a和图1b),PFC1均与距离最近(图1c和图1d)。最后根据分子和形态学鉴定结果,两种野生植物BQ1和PFC1分别被鉴定为野荸荠()和五棱飘拂草()。
2.2 不同植物根际和非根际土壤化学性质比较
同一种植物的根际与非根际土壤pH无显著差异,但是水稻根际pH显著高于野生植物根际和非根际土壤pH;土壤交换性铝和有效性锰含量在根际与非根际以及不同植物之间均无显著差异(表1)。虽然在水稻种植前采集的未淹水土壤(干土)pH为3.69(见材料与方法),但是在淹水条件下采集的植物根际和非根际土壤pH却均高于4.2(表1)。此外,测定的稻田田面水pH仅有2.0左右。在水稻种植中,淹水条件下土壤中较多的H+释放至水体中,导致土壤pH升高,而水体pH急剧降低,这种强酸性的水体环境可能会严重抑制水稻生长。
2.3 不同植物根和地上部矿质元素含量比较
根矿质元素含量分析(表2)表明,2种野生植物P、K含量均显著高于水稻,前者是后者的1.82倍~5.54倍;野荸荠Na含量显著高于水稻,五棱飘拂草Na含量在野荸荠和水稻之间;其他7种元素(N、Ca、Mg、S、Al、Fe、Mn)含量在3种植物之间无显著差异。
地上部矿质元素含量分析(表3)表明,N含量在3种植物之间无显著差异;2种野生植物P、K、Ca、Mg、S、Na、Mn含量均显著高于水稻,前者为后者的1.41倍~9.67倍,其中K和Na的倍数最高;水稻地上部Al和Fe含量显著高于野荸荠,与五棱飘拂草之间无显著差异(表3)。这些结果表明,水稻较野生植物更多地吸收了Al和Fe,但是却较少地吸收了养分(N除外)。

表1 不同植物根际和非根际土壤化学性质
注:同一列不同字母表示不同土壤之间在0.05水平上差异显著。Note:Different letters in the same column indicate significant differences between soils at the level of 0.05

表2 不同植物根部矿质元素含量
注:Mn的浓度单位为mg·kg–1,其他元素的浓度单位均为g·kg–1;同一行不同小写字母表示三种植物之间在0.05水平上差异显著。下同。Note:mg·kg–1as unit for Mn concentration,and g·kg–1as unit for other elements;Different letters in the same line indicate significant differences between plants at the level of 0.05. The same below

表3 不同植物地上部矿质元素含量
2.4 不同植物体pH比较
野荸荠地上部pH显著高于水稻与五棱飘拂草,水稻与五棱飘拂草之间地上部pH无显著差异,3种植物根部pH亦无显著差异(表4)。
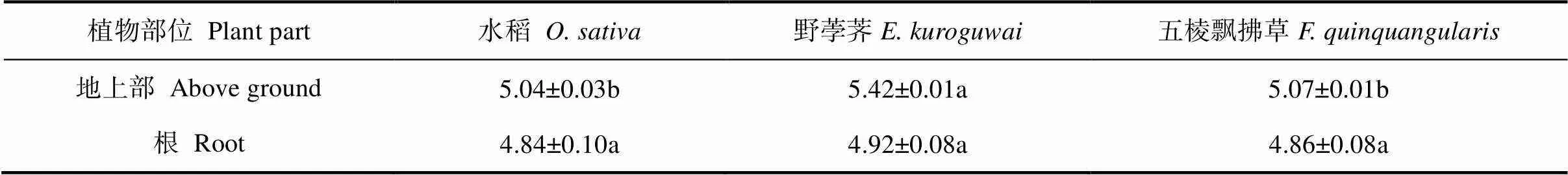
表4 不同植物体pH
注:同一行不同字母表示不同植物之间在0.05水平上差异显著。Note:Different letters in the same line indicate significant differences between plants at the level of 0.05.
3 讨 论
3.1 耐酸性硫酸盐土野生植物资源挖掘
酸性土壤中酸害、铝毒、锰毒、铁毒以及氮、磷、钙、镁缺乏等被认为是限制植物生长的因子[17-20]。大多数粮食作物在酸性土壤上生长均会受到严重抑制,但是一些野生植物却能在酸性土壤上正常生长[14,19],表明野生植物存在一些特殊的耐酸性土壤机制。例如,在对酸性硫酸盐土上野生植物的研究[21]中,飘拂草属()中的两岐飘拂草(subsp)对铝和酸有很强的耐受性,400 mg·L–1Al3+和pH 2.0环境对该植物的生长均无显著影响。相似地,和也在酸性硫酸盐土上被发现[22]。另据前人报道,荸荠属()的多个种(如和)也能在酸性硫酸盐土上正常生长[23-24]。研究发现,适应酸性硫酸盐土的大多数野生植物属于莎草科(),荸荠属和飘拂草属均属于莎草科[22]。本研究也在我国南方酸性硫酸盐土上发现了两种野生植物,即使研究的240个水稻品种均不能完成整个生育期,但是这两种野生植物却能够茂盛地生长。进一步通过形态学和分子鉴定,这两种野生植物被鉴定为野荸荠()和五棱飘拂草(),这为植物耐酸性硫酸盐土机制研究提供了新的植物资源。
3.2 水稻和野生植物对酸性硫酸盐土耐性差异机制
由于野生植物大多不易培养,相关遗传和分子信息不清楚,野生植物的耐酸性土壤机制亦不清楚,或者机制揭示不够深入。特别对于酸性硫酸盐土这种强酸性土壤,生长于其中的野生植物的耐性机制研究更少,大多侧重于报道一些野生植物能够在酸性硫酸盐土上生长,而缺乏对相关机制的探讨,更未见到野生植物和水稻进行比较的相关报道。在本研究中,虽然野荸荠和五棱飘拂草能够在酸性硫酸盐土上良好生长,但是水稻却不能正常生长。与当地农民耕种的正常地块水稻相比,试验地块里的水稻生育期整体延迟,虽然可以完成营养生长期,但是无法完成生殖生长期。
据报道,有些耐铝植物适应酸性土壤的机制是根际碱化,从而降低根际环境里的铝浓度而保持正常生长,且耐铝品种能够更有效地提高根际pH[25-26]。本研究中,无论是水稻还是野生植物,根际和非根际土壤pH均无显著差异(表1),而一般认为根际土和非根际土壤性质存在差异。本文所研究的酸性硫酸盐土极酸,在田间原位监测的稻田田面水pH仅有2.0,植物本身对这种极酸环境下根际土壤pH的改变可能比较小。此外,在这种淹水条件下,由于物质流动性较强,根际和非根际土壤pH之间差异可能比较小。水稻根际土壤pH虽然高于两种野生植物根际土壤pH,但是根际土壤交换性铝和有效锰含量在野生植物和水稻间并无差异,水稻仍然无法像野荸荠和五棱飘拂草一样正常生长。在这种极端土壤逆境环境下,虽然可能由于本能性反应,水稻趋向于通过提高根际土壤pH缓解毒害作用,但是水稻不能通过一般作物调控根际微环境的途径,来完全消除酸性硫酸盐土的强酸害。因此,水稻根际pH较高的效应被大大削弱,根际碱化效应不足以让水稻完全适应这种强酸性土壤。相反,因为两种野生植物本身对酸性硫酸盐土具有高耐性,所以两种野生植物根际pH未升高,结果是两种野生植物根际土壤pH低于水稻。与该结果相似,有文章也报道高耐酸铝的两歧飘拂草()的根际土壤pH显著低于另外一种低耐酸铝芒属植物()[21]。最近一系列研究表明,在水培条件下,培养溶液pH升高反而会增加铝对植物和微生物的毒害[27-28]。根据以上分析,推断野荸荠与五棱飘拂草耐酸性硫酸盐土的机制不是通过诱导根际pH升高,可能是通过自身的其他机制来适应酸性硫酸盐土。在恶劣的酸性硫酸盐土上,一般农作物根系调控的根际微环境变化起到的作用极弱,所以不能正常生长。
虽然水稻根际土壤pH高于野生植物根际和非根际土壤pH(表1),但是水稻地上部较野荸荠地上部积累了更多的铁和铝(表3)。出现这种现象的原因可能是野荸荠本身具有将铁和铝排除在体外的机制,植物积累铝和铁的多少主要取决于植物本身的遗传因素。前人研究[23]表明,木贼状荸荠()是一种非常耐酸性环境的植物,这种植物能将除锰外的一些其他微量金属元素包括铝和铁从植物地上部排除出去,从而更好地适应酸性环境。与这篇报道相类似,本文结果也表明,水稻与野荸荠之间根系铝、铁、锰3种元素含量无差异,但是水稻地上部铝和铁含量显著高于野荸荠,水稻地上部锰含量反而低于野荸荠(表3)。这表明虽然野荸荠根系吸收了大量的铝铁,但是从根系向地上部的转移量较水稻要少,地上部积累较少的铝和铁是野荸荠适应酸性硫酸盐土的主要机制。同时,野荸荠植物体pH显著高于水稻和五棱飘拂草(表4),原因可能是因为野荸荠体内积累的酸性离子铝离子较少,而相应的其他碱性金属元素较多,导致野荸荠地上部pH较高。三种植物根系矿质元素含量相差较小(表2),根系pH亦无显著差异(表4)。对水稻而言,极酸的水体环境和恶劣的土壤性质超出了水稻对逆境的适应能力,致使地上部积累较多的铝和铁,这是水稻不能在酸性硫酸盐土上良好生长的原因。虽然锰毒也是酸性土壤上植物生长的限制因子之一[29],但是由于锰的移动性强,在酸性硫酸盐土壤上有强烈的淋溶作用[30],土壤中有效锰的浓度并不高(表1),且水稻和野荸荠根和地上部锰含量均不高(表2和表3),不足以对植物造成毒害。前人[31]报道也表明酸性硫酸盐土上水稻生长不良的主要原因是铝毒和铁毒,而不是锰毒。因此,目前试验条件下野荸荠和水稻适应酸性硫酸盐土能力的差异是因为铝毒和铁毒,而不是因为锰毒。
与野荸荠相同,五棱飘拂草与水稻之间根系铝和铁含量无显著差异(表2),但是与野荸荠不同的是,五棱飘拂草与水稻之间地上部铝和铁含量亦无显著差异(表3)。这表明五棱飘拂草适应酸性硫酸盐土的机制有别于野荸荠。Yamamoto等[21]对两歧飘拂草()的研究发现,两歧飘拂草能忍耐高浓度铝和极低pH(2.0)环境,并推测其可能有自身的耐铝机制。本文结果表明,虽然五棱飘拂草地上部铝和铁含量略低于水稻,但是二者差异不显著(表3),水稻不能在酸性硫酸盐土上正常生长,而五棱飘拂草可以。与野荸荠通过地上部对铝、铁的排斥机制不同,推测五棱飘拂草对铝、铁的适应机制可能是内部忍耐机制。一些铝超积累植物可以通过内部忍耐机制如液泡分室作用来解铝毒[32]。一般认为水稻耐铝机制以排斥机制为主,使大部分铝集中于水稻根部,而地上部铝含量很低[33]。这里的结果也表明水稻地上部铝含量确实远远低于根铝含量。一般植物地上部铝含量大于1 000 mg·kg–1时,便被定义为铝积累植物[34]。在目前所研究的酸性硫酸盐土上,水稻体内铝含量远超过了1 000 mg·kg–1(表3),这必然对水稻生长产生毒害。水稻积累如此高的铝可能与酸性硫酸盐土的极酸水环境有关,田间原位测定的稻田田面水pH为2.0左右,这必然加剧铝的溶出,进而加剧了水稻地上部铝积累。此外,铁的毒害也是酸性硫酸盐土水稻生长受抑制的一个原因[35]。综合以上信息推断,吸收了过量的铝和铁可能是酸性硫酸盐土上水稻无法正常生长的原因,至于五棱飘拂草的具体内部耐性机制,尚需进一步研究。
虽然水稻地上部积累了较多的铝和铁,但有趣的是,与野荸荠和五棱飘拂草相比,水稻地上部各种矿质养分元素(氮除外)含量却较低(表3)。Osaki等[14]对酸性硫酸盐土上多种野生植物在不同土壤环境下体内铝含量与其他元素含量做了相关性分析,结果表明,植物体内铝含量与多种元素(P、K、Ca、Mg、Fe、Zn、Cu、Na)含量呈负相关,而与氮和锰含量无相关性。这一结果与目前的结果十分相似,即植物吸收的铝越多,吸收的养分越少。与Osaki等[14]报道不同的是,目前的研究表明水稻体内铝多、铁也多,野荸荠体内铝少、铁也少,看起来铝和铁呈正相关,这可能是个特例。虽然Na不是植物必需营养元素,但是Na和K呈现出了相似规律,这可能是因为植物对K和Na的吸收具有相似机制。对于生长在酸性硫酸盐土上的水稻而言,水稻对营养元素的吸收及体内维持较高营养元素含量的能力远远不及野荸荠和五棱飘拂草。以前的报道[17-18]也表明铝毒会抑制植物对多种养分的吸收。笔者推测水稻根系吸收了大量铁、铝,并在地上部积累了高浓度铝和铁,产生了铝毒和铁毒,抑制了根系生长及养分吸收和运输系统,导致水稻吸收和转运养分的能力远远弱于野荸荠和五棱飘拂草,养分缺乏进一步抑制了水稻生长。这从养分吸收角度进一步解释了野生植物和水稻耐酸性硫酸盐土差异的原因。通过调控植物对养分的吸收,可以增强植物抵抗酸性土壤的能力[18]。
3.3 酸性硫酸盐土的利用
酸性硫酸盐土作为水热资源丰富地区的一种土壤类型,应充分发挥其利用价值。通过对水稻与生长无障碍的野生植物对矿质元素吸收进行比较,可以了解酸性硫酸盐土上作物特需的营养元素,进而针对性地对作物进行合理补充养分,进行科学施肥。同时,筛选一些酸铝胁迫下具有较高养分吸收利用效率的水稻品种,增强水稻在酸性硫酸盐土上的栽培和种植技术,可以发挥酸性硫酸盐土上作物的生产潜力。此外,国内外对酸性硫酸盐土上生长的野生植物耐性机制研究较少。野生植物在如此恶劣的土壤上较好地生长,对逆境的抵抗和对营养元素的吸收、利用可能有其特别的机制。研究野生植物在极端酸性环境下对营养元素的高效利用机制,不仅可以为提高酸性硫酸盐土作物生产力提供理论参考,还能为废弃矿区的生物修复提供植物资源和技术。
4 结 论
在所试验的酸性硫酸盐土(pH 3.69)上,种植的240个水稻品种均不能正常生长,却发现两种野生植物可以茂盛地生长。经形态学和分子系统进化树的方法,两种野生植物被鉴定为野荸荠和五棱飘拂草。根际土壤pH、交换性铝含量、有效锰含量均不能解释水稻和两种野生植物适应酸性硫酸盐土能力的差异,即两种野生植物不是通过诱导根际土壤性质变化来适应酸性硫酸盐土。目前试验条件下,锰毒耐性、氮高效吸收与植物适应酸性硫酸盐土能力关联亦较小。水稻地上部积累了较多铝和铁,却积累了较少养分;野荸荠地上部积累了较少铝和铁,却积累了较多养分;五棱飘拂草地上部积累了与水稻相似量的铝和铁,也积累了较多养分。根系各种矿质元素含量在不同植物之间差异较小。根据上述结果,推断铝毒和铁毒及养分吸收能力降低是酸性硫酸盐土上水稻生长的限制因子,而野荸荠和五棱飘拂草因为能够很好地抵抗铝毒和铁毒,养分吸收能力也较强,所以两种野生植物可在酸性硫酸盐土上良好生长。野荸荠可能对铝和铁存在排斥机制,限制铝和铁向地上部转运,而五棱飘拂草对铝和铁的内部忍耐能力较强,能在体内积累高含量的铝和铁,却不受毒害。
致 谢 感谢南京农业大学万建民教授和江玲教授提供水稻种质资源。
[1] Powell B,Martens M. A review of acid sulfate soil impacts,actions and policies that impact on water quality in Great Barrier Reef catchments,including a case study on remediation at East Trinity. Marine Pollution Bulletin,2005,51(1):149—164.
[2] Dent D L,Pons L J. A world perspective on acid sulphate soils. Geoderma,1995,67(3):263—276.
[3] Ljung K,Maley F,Cook A,et al. Acid sulfate soils and human health-a millennium ecosystem assessment. Environment International,2009,35(8):1234—1242.
[4] Huang Q Y,Tang S H,Lu Y,et al. Formation,characteristics and ecological effects of acid sulfate soils. Plant Nutrition and Fertilizer Science,2014,20(6):1534—1544. [黄巧义,唐拴虎,卢瑛,等. 酸性硫酸盐土的形成、特性及其生态环境效应. 植物营养与肥料学报,2014,20(6):1534—1544.]
[5] Gong Z T,Zhang X P. Mangrove and acid sulphate soils in China. Acta Pedologica Sinica,1994,31(1):86—94. [龚子同,张效朴. 中国的红树林与酸性硫酸盐土. 土壤学报,1994,31(1):86—94.]
[6] Zhao Q G,Yang J S,Zhou H.“Ten words”strategic policy for ensuring red line of farmland and food security in China. Soils,2011,43(5):681—687. [赵其国,杨劲松,周华. 保障我国“耕地红线”及“粮食安全”十字战略方针. 土壤,2011,43(5):681—687.]
[7] Zhang J E. The process of acid hazards in acid sulfate soils and its environmental effects. Tropical Geography,1999,19(2):137—141. [章家恩. 酸性硫酸盐土的酸害暴发机制及其环境影响. 热带地理,1999,19(2):137—141.]
[8] Huang J M. Chemical characteristics of strong acidic saline paddy soils(acid-reversing fields)in Guangdong coastal waters. Acta Pedologica Sinica,1958,6(2):114—122. [黄继茂. 广东滨海强酸性盐渍水稻土(反酸田)化学特性的研究. 土壤学报,1958,6(2):114—122.]
[9] Huang Q Y,Tang S H,Huang X,et al. Analysis on the limiting hazardous chemical factors of acid sulfate paddy soils in Guangdong Province. Plant Nutrition and Fertilizer Science,2016,22(1):180—191. [黄巧义,唐拴虎,黄旭,等. 广东省酸性硫酸盐水稻土作物产量的主要限制因子分析. 植物营养与肥料学报,2016,22(1):180—191.]
[10] Zhang J E. The landscape diagnosis and their spatial and temporal patterns of distribution and succession of sulphate acid soils. Soil and Environmental Sciences,1999,8(2):144—147. [章家恩. 酸性硫酸盐土的景观诊断及时空分布与演替模式. 土壤与环境,1999,8(2):144—147.]
[11] Zang X P,Zhang C L,Sun G M,et al. Effect of phosphate rocks on nutrient availability of rice on acid sulfate soils. Plant Nutrition and Fertilizer Science,2003,9(2):203—207. [臧小平,张承林,孙光明,等. 酸性硫酸盐土壤上施用磷矿粉对水稻养分有效性的影响. 植物营养与肥料学报,2003,9(2):203—207.]
[12] Liu Z Q,Duan S S,Li A F,et al. Transformation characteristics of sulfur forms in acid sulfate soils under different soil water conditions. Chinese Journal of Applied Ecology,2004,15(9):1570—1574. [刘振乾,段舜山,李爱芬,等. 不同土壤水分条件下酸性硫酸盐土硫形态转化特征. 应用生态学报,2004,15(9):1570—1574.]
[13] Liu Z Q,Wang J W,Luo S M,et al. Hydrodynamic mechanism on the change of different forms of acid in acid sulfate soil. Acta Pedologica Sinica,2002,39(5):726—734. [刘振乾,王建武,骆世明,等. 酸性硫酸盐土酸消长的水动力机制研究. 土壤学报,2002,39(5):726—734.]
[14] Osaki M,Watanabe T,Ishizawa T,et al. Nutritional characteristics in leaves of native plants grown in acid sulfate,peat,sandy podzolic,and saline soils distributed in Peninsular Thailand. Plant and Soil,1998,201(2):175—182.
[15] Wang Y H. Food analysis. Beijing:Chemical Industry Press,2010. [王永华. 食品分析. 北京:化学工业出版社,2010.]
[16] Chen S L,Yao H,Han J P,et al. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS One,2010,5(1):e8613.
[17] Zhao X Q,Chen R F,Shen R F. Co-adaptation of plants to multiple stresses in acidic soils. Soil Science,2014,179(10/11):503—513.
[18] Zhao X Q,Shen R F. Strategies for increasing the utilization of nitrogen and phosphorus by plants under aluminum stress. Plant Physiology Journal,2015,51(10):1583—1589. [赵学强,沈仁芳. 提高铝毒胁迫下植物氮磷利用的策略分析. 植物生理学报,2015,51(10):1583—1589.]
[19] Chen R F,Dong X Y,Zhao X Q,et al. Advances in adaptive mechanisms to acid soils in woody plants—with an emphasis on()and oil tea(). Soils,2015,47(2):252—258. [陈荣府,董晓英,赵学强,等. 木本植物适应酸性土壤机理的研究进展——以胡枝子()和油茶()为例. 土壤,2015,47(2):252—258.]
[20] Moore P A,Attanandana T,Patrick W H. Factors affecting rice growth on acid sulfate soils. Soil Science Society of America Journal,1990,54(6):1651—1656.
[21] Yamamoto A,Wasaki J,Funatsu Y,et al. Distribution and stress tolerance ofsubsp()growing in highly acidic solfatara fields. Ecological Research,2018,33(5):971—978.
[22] Uchiyama H,Matoba H,Aizawa T,et al. Chromosome counts of some wetlandspecies from the Mekong Delta,Vietnam. Cytologia,2010,75(4):335—339.
[23] Lottermoser B G,Ashley P M. Trace element uptake by(spike rush)in an abandoned acid mine tailings pond,northeastern Australia:implications for land and water reclamation in tropical regions. Environmental Pollution,2011,159(10):3028—3035.
[24] Aizawa T,Ve N B,Nakajima M,et al.sp. nov.,a nitrogen-fixing bacterium isolated from an aquatic plant,,that grows in highly acidic swamps in actual acid sulfate soil areas of Vietnam. International Journal of Systematic and Evolutionary Microbiology,2010,60(5):1152—1157.
[25] Foy C D,Burns G R,Brown J C,et al. Differential aluminum tolerance of two wheat varieties associated with plant-induced pH changes around their roots. Soil Science Society of America Journal,1965,29(1):64—67.
[26] Degenhardt J,Larsen P B,Howell S H,et al. Aluminum resistance in the Arabidopsis mutantis caused by an aluminum-induced increase in rhizosphere pH. Plant Physiology,1998,117(1):19—27.
[27] Zhao X Q,Bao X M,Wang C,et al. Hydroxy-Al and cell-surface negativity are responsible for the enhanced sensitivity ofto aluminum by increased medium pH. Archives of Microbiology,2017,199(8):1185—1194.
[28] Wang W,Zhao X Q,Chen R F,et al. Altered cell wall properties are responsible for ammonium-reduced aluminum accumulation in rice roots. Plant,Cell and Environment,2015,38(7):1382—1390.
[29] EI-Jaoual T,Cox D A. Manganese toxicity in plants. Journal of Plant Nutrition,1998,21(2):353—386.
[30] Liu Z H,Wang Z Q. Transformation and movement of iron and manganese in acid sulphate soils in China. Acta Pedologica Sinica,1994,31(4):376—384. [刘兆辉,王遵亲. 我国酸性硫酸盐土壤中铁锰形态转化及迁移. 土壤学报,1994,31(4):376—384.]
[31] Kang D J,Seo Y J,Lee B K,et al. Identification and crop performance of acid sulfate soil-tolerant rice varieties. Journal of Crop Science and Biotechnology,2010,13(2):75—81.
[32] Shen R F,Ma J F,Kyo M,et al. Compartmentation of aluminium in leaves of an Al-accumulator,Moench. Planta,2002,215(3):394—398.
[33] Chen RF,Shen RF,Gu P,et al. Response of rice()with root surface iron plaque under aluminium stress. Annals of Botany,2006,98(2):389—395.
[34] Chenery E M,Sporne K R. A note on the evolutionary status of aluminium-accumulators among dicotyledons. New Phytologist,1976,76(3):551—554.
[35] Sahrawat K L. Iron toxicity to rice in an acid sulfate soil as influenced by water regimes. Plant and Soil,1979,51(1):143—144.
Comparison of Rice Plants with Two Wild Plants in Tolerance to Acid Sulfate Soil and Absorption of Mineral Elements
ZHANG Lingyu1,3, ZHAO Xueqiang1,3†, LI Jiamei2, SHEN Renfang1,3
(1. State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China; 2. College of Life Science, Henan Agricultural University, Zhengzhou 450002, China; 3. University of Chinese Academy of Sciences, Beijing 100049, China)
Acid sulfate soil is a type of soil developed from the parent material of reducing sulfide and is described as “the nastiest soils in the world” because of its strong acidity and poor plant growth. Acid sulfate soils are mainly distributed along the seacoast of tropical and subtropical regions, wherein the climate is though very suitable for growth of most crops, especially rice (L.) because of their abundant water and heat resources, it is very hard to bring the production potential of rice into full play, due to their poor soil properties, such as acid threats, metal toxicity and nutrient deficiency. Plants, the same in family, but different in species or variety, vary significantly in capability to adapt to stress soils. Although most crop plants cannot grow in acid sulfate soils, some native plants can do very well. The objective of this study is to screen out rice cultivars that are capable to adapt to acid sulfate soils, and further to explore their underlying mechanisms.A total of 240 varieties of rice were planted in acid sulfate soil (pH 3.69) in Taishan City, Guangdong Province, China, and were managed as the local farmers do their crops. None met the target of economic yield. However, accidentally two species of wild plants, identified morphologically and molecularly asandwere found growing flourishingly in the soil. During the rice harvest season, samples of the rice plants, wild plants, and their rhizosphere and non-rhizosphere soils were collected for analysis of soil chemical properties (pH, exchangeable Al, and available Mn) and contents of 10 elements (N, P, K, Ca, Mg, S, Na, Al, Fe, and Mn) in the roots and shoots.It was found that variation of soil chemical properties such as pH, exchangeable Al and available Mn, in rhizosphere could not explain the difference between the two wild plants and the rice plants in tolerance and that Mn toxicity and N deficiency were not the key factors that limited growth of the plants. Instead, the contents of mineral elements in the plant shoots were found to be closely related to the difference between the rice plants and the wild plants in tolerance. The rice plants accumulated more Al and Fe, but less nutrients (P, K, Ca, Mg, and S) in the shoots than the two wild plants. Little difference was observed in contents of most mineral elements in roots between the plants. Moreover, the Al and Fe contents in the shoot ofwere low, which explains its tolerance to Al and Fe toxicity through exclusion mechanism, while the Al and Fe contents in the shoot ofwere high, which explains its resistance to Al and Fe toxicity through internal tolerance mechanism.Al toxicity, Fe toxicity and poor nutrient uptake are the main factors leading to poor growth of rice in acid sulfate soils. In contrast, the two wild plantsandcan grow well in the soils thanks to their strong ability to resist Al and Fe toxicity and to take up nutrients. Mn toxicity and N deficiency may not be involved in the difference between the rice plants and the wild plants in adaptability to acid sulfate soils. It is, therefore, suggested that to improve Al and Fe tolerance and nutrient uptake are two approaches key to developing crop productivity potential in acid sulfate soils, which can be achieved through soil management and variety breeding.
Acid sulfate soil; Rice;;; Metal toxicity; Nutrient deficiency
S158
A
10.11766/trxb201901300064
张玲玉,赵学强,李家美,沈仁芳. 水稻和两种野生植物对酸性硫酸盐土耐性及矿质元素吸收[J]. 土壤学报,2020,57(2):403–413.
ZHANG Lingyu,ZHAO Xueqiang,LI Jiamei,SHEN Renfang. Comparison of Rice Plants with Two Wild Plants in Tolerance to Acid Sulfate Soil and Absorption of Mineral Elements [J]. Acta Pedologica Sinica,2020,57(2):403–413.
* 国家自然科学基金项目(31672229)和中国科学院重点部署项目(KFZD-SW-112-01-09)共同资助 Supported by the National Natural Science Foundation of China(No. 31672229)and the Key Deployment Projects of the Chinese Academy of Sciences(No. KFZD-SW-112-01-09)
,E-mail:xqzhao@issas.ac.cn
张玲玉(1992—),女,山东淄博人,硕士研究生,主要从事酸性土壤植物营养研究。E-mail: zhangly@issas.ac.cn
2019–01–30;
2019–03–19;
优先数字出版日期(www.cnki.net):2019–04–15
(责任编辑:陈荣府)
