伴生细菌在入侵种桉树枝瘿姬小蜂克服桉树抗性中的作用
2020-04-23寇冀蒙刘芳华刘一澎
寇冀蒙, 刘芳华, 刘一澎, 马 玲, 鲁 敏
1东北林业大学林学院,黑龙江 哈尔滨 150040; 2中国科学院动物研究所,北京 100101; 3安徽大学物质科学与信息技术研究院,安徽 合肥 230039; 4中国农业科学院植物保护研究所,北京 100193
桉树Eucalyptus属桃金娘科Myrtaceae,原产于澳大利亚和印度尼西亚及其附近岛屿,是世界上著名的速生丰产树种之一(祁述雄,2002)。我国自1890年起先后引进300多个树种(含变种和杂交种),广泛分布在全国的20多个省市以及600多个自治区、县,总种植面积170万hm2(王豁然等,1999; 徐建民等,2001)。桉树枝瘿姬小蜂LeptocybeinvasaFisher & La Salle是近年来新发现的一种主要危害桉属树种外来有害生物,隶属于膜翅目Hymenoptera小蜂总科Chalcidoidea姬小蜂科Eulophidae啮小蜂属Tetrastichinae,原产于大洋洲的澳大利亚昆士兰州(Aytar,2006)。2007年桉树枝瘿姬小蜂首次扩散到中国,并给广西壮族自治区东兴县的桉树资源造成了危害,随后该虫开始陆续出现在广西、广东、海南、福建、江西、湖南、云南等省(自治区)的桉树商品林区,严重影响了桉树产业的正常发展,仅广西,从2007年首次发现至2009年就有6500 hm2桉树受害(常润磊和周旭东,2010)。
植物在面对昆虫进攻时,可以通过很多防御手段对其抵抗。植物的抗虫性可分为组成抗性和诱导抗性。前者是植物的固有特性,因基因型的不同而有差别,始终存在于植物体内并起作用,但会因环境条件的改变而改变抗性程度;而后者则是在遇到外界伤害,如机械损伤、植食者取食和病原菌侵染时植物影响植食者行为或降低其嗜好性的反应(Kangetal.,2018a; Thiboutetal.,1993),是一种类似于免疫反应的抗性现象(Agrawaletal.,1999)。虽然植物具有强大复杂的防御体系,但是植食性昆虫在与植物的长期协同进化中演化出多种反防御措施来维持种群的发展。除了在同种昆虫不同个体间的协同作用中出现外,植食性昆虫的反防御还出现在不同种昆虫之间的交互作用中(Bruinsma & Dicke,2008; Kangetal.,2018b; Kessleretal.,2004),包括行为防御(Bernay,1998)、生理防御与生物化学防御机制等(刘芳华等,2017; Qngsmaetal.,1995; Zhuetal.,2005)。昆虫伴生细菌是指栖息在昆虫肠道、体表、口腔等部位的一些与昆虫关系密切的微生物,它们在昆虫的生活史中扮演着重要的角色(Engel & Moran,2013)。如红脂大小蠹肠道细菌可以帮助昆虫降解寄主油松的单萜(Xuetal.,2016),肠道细菌可以促进玉米根虫对植物的适应力(Chuetal.,2013)。
近年来,有关桉树与桉树枝瘿姬小蜂的互作研究取得了一些突破性进展,但主要集中在桉树抗虫生理生化机理方面(刘慧清,2012; 吕文玲等,2012; 王伟等,2012; 吴耀军等,2010)。这些研究结果表明,在遭受桉树枝瘿姬小蜂危害后,桉树能够通过大量合成植物防御性次生代谢物和激活防御酶活性来抵御桉树枝瘿姬小蜂的持续危害。而桉树枝瘿姬小蜂如何克服寄主桉树抗性的研究还未见报道。
现已报道的植食性昆虫克服寄主抗性主要集中在以下2个方面:自身解毒酶系统和伴生微生物代谢系统(Jeschkeetal.,2016; Luetal.,2016; Sunetal.,2013; Zouetal.,2016)。本研究拟测定桉树次生代谢物质黄酮和单宁的含量,研究分离得到的桉树枝瘿姬小蜂伴生细菌对桉树抗性物质的耐受性和代谢能力进行分析,旨在揭示桉树枝瘿姬小蜂伴生细菌在其寄主定殖过程中是否有协同作用,是否可帮助其寄主克服桉树次生代谢物质的毒害作用,进而为桉树枝瘿姬小蜂的微生物生态研究奠定基础。
1 材料与方法
1.1 供试植物和细菌
供试桉树:DH201-2(巨园桉Eucalyptusgrandis×E.tereticornis)和A107(E.urophylla)来自广西林业科学研究院和广西东口林场。
供试细菌:B1(Staphylococcuscohnii)、B2(Curtobacteriumoceanosedimentum)、B3(Bacilluswiedannii)、B4(Peseudomonasgeniculate)、B5(Serratiamacescens)和B6(Klebsiellaquasipneumoniae)来自中国科学院动物研究所森林化学生态实验室。
1.2 桉树次生代谢物质黄酮和单宁的含量测定
1.2.1 黄酮含量测定 选取不同品系和易感品系危害前后的桉树枝条,烘干至恒重,粉碎过40目筛备用。
测定方法:使用植物类黄酮含量检测试剂盒(Solarbio, China)测定桉树枝条中黄酮的含量。
1.2.2 单宁含量测定 选取不同品系和易感品系危害前后的桉树枝条,烘干至恒重,粉碎过60目筛备用。
测定方法:称取样品0.05 g (3个重复),放入5 mL离心管内,加水4 mL,摇匀,放置在80 ℃水浴上提取30 min,每隔约10 min摇匀一次。然后迅速置于冷水浴中冷却,加水定容到5 mL,摇匀,过滤。若滤液浑浊,重复过滤一次。吸取滤液0.1 mL到试管中,加入4.9 mL的水稀释,然后加入0.2 mL福林酚试剂,摇匀,5 min后加1 mL的饱和碳酸钠溶液,摇匀,此时溶液显示蓝色。室温静置30 min,用酶标仪于760 nm处测定其光密度。同时以蒸馏水作空白调零。
1.3 黄酮和单宁对桉树枝瘿姬小蜂伴生细菌的生长影响
1.3.1 黄酮和单宁对伴生细菌的毒性 黄酮和单宁最低抑菌浓度(minimum inhibitory concentration, MIC)测定实验用来检验次生代谢物质黄酮和单宁是否对桉树枝瘿姬小蜂伴生细菌有利。2种化学物质的最低抑菌浓度实验参考Cosentinoetal. (1999)的培养基液体微稀释法。具体操作如下:(1)所有待测的6株细菌在LB培养基中过夜培养;(2)将黄酮和单宁分别溶解于二甲基亚砜和无菌水中,并加到含有LB培养基的96孔板(Greiner, Germany)中,黄酮按照20、40、60 、80、100、120、140、160 ng·μL-1的浓度稀释,单宁按照500、1000、1500、2000、3000、4000、5000、6000 ng·μL-1的浓度稀释;(3)过夜培养的6株细菌调整其D600 nm为 0.5,并稀释100倍于96孔板中;(4)将96孔板放于25 ℃恒温培养箱中培养12 h;(5)12 h后,检查每个孔,看是否有菌长出(裸眼观察),可以抑制细菌生长的浓度为最低抑菌浓度。以上每种细菌的每个浓度做3个重复。
1.3.2 黄酮和单宁对伴生细菌的生长影响 黄酮对伴生细菌的生长影响:2种细菌(B1和B4)在MRS肉汤培养基上培养。首先,30 ℃恒温摇床180 r·min-1条件下培养24 h,调整细胞密度D600 nm=0.5左右,并将菌液稀释100倍至4 mL MRS肉汤培养基。然后将黄酮溶解于二甲基亚砜(dimethyl sulfoxide, DMSO)中,每只摇菌管中加入40 μL黄酮/DMSO溶液于培养基内,最终使微生物悬浮液浓度分别为10、30、50、60、70、80 ng·μL-1,每一浓度设置5个重复。然后在30 ℃恒温摇床180 r·min-1条件下培养24 h。之后在600 nm可见光条件下检测不同黄酮浓度条件下细菌悬浮液的光密度。对照组中不含黄酮,只含有等量的DMSO,每种细菌都设置一个对照,对照也设置5个重复。
单宁对伴生细菌的生长影响:3种细菌(B3、B5和B6)在M9矿物质盐培养基上培养。首先,30 ℃恒温摇床180 r·min-1条件下培养24 h,调整细胞密度D600 nm=0.5左右,将菌液稀释100倍至4 mL M9矿物质盐培养基。然后将单宁溶解于无菌水中,每只摇菌管中加入40 μL单宁水溶液于培养基内,使微生物悬浮液浓度分别为1000、2000、3000、4000、5000、6000 ng·μL-1,每一浓度设置5个重复。在30 ℃恒温摇床180 r·min-1条件下培养24 h。在600 nm可见光条件下检测不同单宁浓度条件下细菌悬浮液的光密度。对照组中不含单宁,每种细菌都设置一个对照,对照也设置5个重复。
1.4 桉树枝瘿姬小蜂伴生细菌对黄酮和单宁的降解能力测定
1.4.1 伴生细菌对黄酮的降解能力 黄酮降解实验在14 mL摇菌管内进行。向每只摇菌管内加入4.9 mL的MRS肉汤培养基,加入50 μL的黄酮/DMSO溶液(终浓度为50 ng·μL-1),各用50 μL的细菌B1和B4接种,37 ℃恒温摇床180 r·min-1恒温摇床培养。在0、2、12和24 h分别吸取混合液400 μL,12000g离心5 min,弃上清,用400 μL的甲醇溶解沉淀,涡旋混匀后,12000g离心5 min,取上清液100 μL于样品瓶内,储存于-20 ℃用于HPLC上机检测。每种细菌设置6组重复,空白对照未接种细菌,只加入黄酮/DMSO溶液,也设置6组重复。使用Agilent 1100 series 高效液相色谱仪检测样品中黄酮的含量。具体方法如下:流动相A色谱级甲醇;流动相B2‰乙酸水溶液。流动相以 0.8 mL·min-1的流速运行,20 min甲醇上升至5%~30%,5 min内甲醇上升至30%~50%,5 min内甲醇上升至50%~65%,65%的甲醇保持5 min,甲醇在7 min内上升至65%~100%。色谱分离柱为Agilent C30柱(250 min×4.6 mmL.D.),每次进样20 μL,每次在样品检测前先运行同样体积的标准品,做出标准曲线用于计算样品浓度。
1.4.2 伴生细菌对单宁的降解能力 单宁降解实验在摇菌管内进行。向每只摇菌管中加入4.9 mL的M9矿物盐培养基,加入50 μL的单宁水溶液(终浓度为1000 ng·μL-1),各用50 μL的细菌B3、B5和B6接种后,28 ℃恒温摇床180 r·min-1培养。在0、12、24和48 h各吸取混合液1 mL,加入2.4 mL乙醇洗涤,手动摇匀5 min,然后5000 r·min-1离心10 min,重复洗涤一次,将2次提取液混合在一起,放入超声波浴中30 min,充分溶解沉淀,吸取上清液于样品瓶中储存于-20 ℃用于HPLC上机检测。每种细菌设置6组重复,对照组只加入单宁,不接菌,也设置6组重复。使用Agilent 1100 series高效液相色谱仪检测样品中单宁的含量。具体方法如下:流动相A2‰乙酸水溶液;流动相B(超纯水/乙腈/乙酸,78/20/2,V/V/V);流动相以1 mL·min-1的流速运行,0~55 min内乙腈/乙酸水溶液增加至80%,55~57 min内乙腈/乙酸水溶液增加至80%~90%,55~57 min内乙腈/乙酸水溶液保持90%,70~80 min内乙腈/乙酸水溶液增加至90%~95%,甲醇在10 min内增加至95%~100%,90~120 min使用甲醇清洗。色谱分离柱为Agilent C30柱(250 min×4.6 mmL.D.),每次样品进样20 μL,每次在样品检测前先运行同样体积的标准品,做出标准曲线用于计算样品浓度。
2 结果与分析
2.1 桉树次生代谢物质黄酮和单宁的含量
实验结果显示,不同品系(A107和DH201-2)健康的桉树次生代谢物质黄酮和单宁的含量与高感品系(DH201-2)桉树遭受小蜂危害后黄酮和单宁的含量差异显著。在高抗品系A107中类黄酮和单宁含量均显著高于高感品系DH201-2(独立样本t检验,P<0.001)。其中,A107中黄酮、单宁含量分别为(4.32±0.12)、(62.62±1.57) mg·g-1;DH201-2中黄酮、单宁含量分别为(1.43±0.23)、(45.75±2.42) mg·g-1。高感品系桉树DH201-2在遭受桉树枝瘿姬小蜂危害前后体内黄酮和单宁含量发生明显变化,危害后黄酮、单宁含量分别为(4.11±0.07)、(82.06±1.76) mg·g-1。
2.2 桉树次生代谢物质黄酮和单宁对伴生细菌的毒性
最低抑菌实验结果显示,黄酮的抑菌范围从40 ng·μL-1到80 ng·μL-1,而单宁的抑菌范围则从1000 ng·μL-1到6000 ng·μL-1(表1和表2)。6株细菌对黄酮和单宁的耐受存在差异:B2、B3、B5和B6对黄酮比较敏感,而B1和B4对黄酮耐受能力较强;B1、B2和B4对单宁较为敏感,而B3、B5和B6对单宁耐受能力较强。

表1 6株伴生细菌对黄酮和单宁的最低抑菌浓度(ng·μL-1)

表2 6株伴生细菌对黄酮和单宁的最低抑菌浓度(ng·μL-1)的描述性统计
2.3 桉树次生代谢物质黄酮和单宁对伴生细菌的生长影响
7种浓度0、10、30、50、60、70和80 ng·μL-1的黄酮对桉树枝瘿姬小蜂伴生细菌B1和B4的生长影响见图1A。其中,10 ng·μL-1的黄酮对2种细菌均无明显的抑制作用,在其余浓度下,随着黄酮浓度的升高,其对细菌的抑制作用越明显。在高浓度80 ng·μL-1黄酮的条件下,2种细菌生长受到完全抑制。
7种浓度0、1000、2000、3000、4000、5000和6000 ng·μL-1的单宁对桉树枝瘿姬小蜂伴生细菌B3、B5和B6的生长影响见图1B。在低浓度1000 ng·μL-1单宁条件下,3种细菌生长均不受影响;在单宁浓度超过2000 ng·μL-1开始,各种细菌的生长均会受到抑制,细菌B3在超过5000 ng·μL-1单宁条件下则完全不能生长,细菌B6在6000 ng·μL-1单宁条件下生长受到完全抑制。
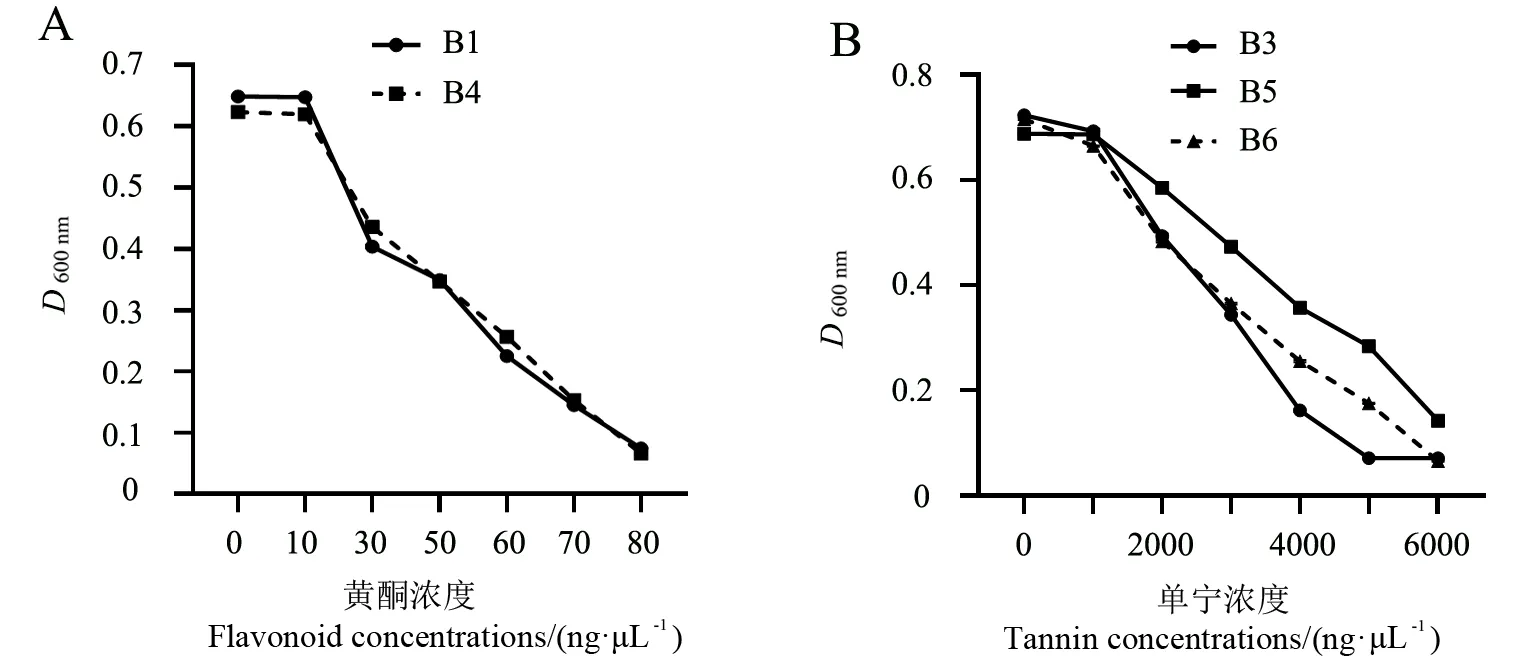
图1 伴生细菌在不同浓度黄酮和单宁条件下的生长情况
2.4 伴生细菌对黄酮和单宁的降解能力
2种细菌B1和B4在不同时间降解黄酮的能力见图2A。其中,在24 h处2种细菌对黄酮的降解率分别达到了27.35%和21.14%,说明2种细菌具有一定的降解黄酮的能力。
3种细菌B3、B5和B6在不同时间降解单宁的能力见图2B。其中,B6在12、24和48 h单宁浓度没有显著差异,说明该细菌对单宁降解能力一般。而B3、B5在48 h时单宁的降解率分别达到了44.82%和48.36%,2种细菌均具有较强的降解单宁的能力(单因素方差分析,P<0.05)。

图2 伴生细菌在不同时间降解黄酮和单宁的浓度变化
3 讨论
在植物与昆虫的长期协同进化过程中,植物为避免植食性昆虫的危害,形成或诱导产生多种特有的次生化合物,这些化合物对大多数昆虫起着化学防御作用(冼继东等,2003)。这些植物次生物质虽非营养物质,但具有防御和保护植物免受昆虫危害的功能。有研究表明,植物在抵御病虫侵害中,次生代谢产物比任何其他自然因素都更为重要和有效。随着植食性昆虫的取食,许多次生代谢产物在昆虫体内不断积累,从而使昆虫生长发育减缓或繁殖率降低,甚至导致昆虫死亡(吕蔷,2010; 薛英伟等,2009; 张彦广等,2001; 周丹丹,2008)。桉叶中黄酮类化合物的研究较早,20世纪60年代Hillis (1966)研究了200余种桉叶中的黄酮类化合物,发现大多含有槲皮素、杨梅素和山奈酚。近年来,研究者从桉属植物中得到了38个黄酮类化合物,其结构为黄酮或黄酮醇及其苷类、二氢黄酮或者二氢黄酮醇及其苷类(Amakuraetal.,2009)。Houetal. (2000)从桉叶中分离出一种没食子单宁酸成分和4种可水解的单宁酸、没食子酸及儿茶酚等。张华峰(2013)发现,桉树枝瘿姬小蜂产卵能明显影响桉树叶黄酮、单宁和总酚等次生代谢物质的含量。而戴沿海(2006)也发现,松树通过增加黄酮的合成量来抵御松突圆蚧的危害。此外,落叶松针叶中单宁等物质含量与松毛虫危害程度呈负相关,说明了单宁是一类可作为树种抗性的标志的化学防御物质(王燕等,2001)。石媛媛等(2017)通过对混交林和纯林中油松不同处理,发现松毛虫取食可诱导增加油松针叶内的缩合单宁和黄酮含量,进而增强油松的抗虫性。本研究发现,高抗品系和被桉树枝瘿姬小蜂危害后的高感品系中黄酮和单宁的含量显著性高于健康高感品系。以上结果表明,黄酮和单宁是桉树低于桉树枝瘿姬小蜂危害的主要防御性代谢物质。
在植物与植食性昆虫协同进化过程中,植物在不断完善其防御反应,同时植食性昆虫也在选择压下不断适应植物防御反应。在植食性昆虫适应植物防御反应的反防御策略也呈现多样性。如昆虫利用其唾液及伴生微生物抑制植物防御反应和降解其次生代谢物等(Engel & Moran,2013; Hammer & Bowers,2015; Hansen & Woran,2014;Linetal.,2015; Liuetal.,2016; Masonetal.,2014)。黄酮类化合物经人体摄入后,大部分不经降解而进入大肠,经由肠道菌群酶的生物转化成简单酚酸后被吸收或排泄(Serraetal.,2012)。周锡钦等(2009)研究发现,从黄芩中提取的黄酮类化合物能够显著抑制白色念珠菌Moniliaalbican的生长。黄酮类化合物对金黄色葡萄球菌Staphylococcusaureus、大肠埃希杆菌Escherichiacoli和枯草芽孢杆菌Bacillussubtilis等的抑制作用较为明显。单宁能够抑制多种革兰氏阳性菌(如金黄色葡萄球菌、链球菌等)和革兰氏阴性菌(如沙门菌、螺杆菌、大肠杆菌、梭菌、弯曲杆菌和芽孢杆菌等)的生长,并且革兰氏阳性菌通常比革兰氏阴性菌更敏感(Daglia,2012)。黄文和石碧(2003)和吕远平等(2003)分别从堆放橡椀的土壤中分离到能降解单宁的黑曲霉、内胞霉和假丝酵母。黄文等(2003)从内胞霉菌丝体中不但检测出单宁酶活性,还检测到多酚氧化酶(PPO)活性,并证实单宁的降解不仅与单宁酶有关,还与多酚氧化酶有关。本研究发现,一些桉树枝瘿姬小蜂伴生细菌具有对黄酮和单宁的高耐受性和代谢能力。进而推测其可能在桉树枝瘿姬小蜂克服桉树抗性的过程中扮演了重要作用。但其在自然环境中的实际能力还需后续研究证明,同时其降解的机制也值得进一步研究。
综上所述,本研究发现高抗品系(A107)桉树体内次生代谢物质黄酮和单宁的含量均显著性高于高感品系(DH201-2)桉树。同时,高感品系桉树遭受桉树枝瘿姬小蜂危害后,体内次生代谢物质黄酮和单宁的含量显著提高,2种物质是桉树抵御桉树枝瘿姬小蜂侵害的主要防御物质,说明桉树枝瘿姬小蜂诱导桉树产生了抗性,高浓度的黄酮和单宁会抑制桉树枝瘿姬小蜂伴生细菌的生长。在低浓度情况下,伴生细菌可以适应,并继续繁殖;在高浓度情况下,主要伴生细菌会被毒杀,而且黄酮的抑制作用更为明显。桉树枝瘿姬小蜂主要伴生细菌在一定浓度的次生代谢产物黄酮和单宁条件下能够生存,并且具有降解黄酮和单宁的能力。在一定的时间内,可以降解大部分次生代谢物质,帮助宿主桉树枝瘿姬小蜂解除和适应桉树的黄酮和单宁防御。
