江西南昌梅岭景区蜘蛛多样性调查及成因分析
2020-04-22陈连水袁凤辉周谷春杜世胜张燕艳
陈连水,袁凤辉,周谷春,饶 军,杜世胜,张燕艳
江西南昌梅岭景区蜘蛛多样性调查及成因分析
陈连水1,袁凤辉1,周谷春2,饶 军1,杜世胜1,张燕艳1
(1. 东华理工大学 生物系,江西 南昌 330013;2. 赣南师范大学 生命科学学院,江西 赣州 341000)
在对江西南昌市梅岭景区6个典型景点中的蜘蛛多样性进行调查、鉴定并获得其蜘蛛资源详实分布名录的基础上,结合另文对其多样性的比较研究结果,通过将其39个未确定种的各科属级物种丰富度、景点物种多样性、群落结构多样性和捕食功能多样性共13个指数作为变量对其区域特色种作用因子进行主成分、相关性和聚类等统计分析,揭示其多样性形成原因和方式。结果表明:景点物种均匀度、景点物种丰富度、景点物种优势度、捕食功能均匀度、群落结构丰富度、捕食功能丰富度是区域特色种形成的6个主要作用因子,各因子间普遍相关,且生态和遗传因素各有侧重地作用于区域特色种的形成。
南昌;梅岭景区;蜘蛛;多样性;成因
蜘蛛隶属节肢动物门(Arthropoda)蛛形纲(Arachnida)蜘蛛目(Araneae),它具有分布广、种类多、数量大、捕食能力强、食量大、繁殖快、寿命长、居留较稳定等特点,已成为农林生态系统中抑制害虫的重要组成部分[1-2]。据报道,全世界有蜘蛛120科、4 149属、48 306种[3];我国有蜘蛛69科、734属、4 228种[4];江西省有蜘蛛41科、247属、729种[5];有研究表明,蜘蛛的地理分布格局有别于依据哺乳动物建立的世界动物地理区划[6]。因为蜘蛛是植物与昆虫食物链或网中的重要节点[7],所以,研究蜘蛛的多样性对于揭示农林生态系统结构的稳定性和能量与物质循环具有重要意义[8]。另外,蜘蛛具有很强的环境指示作用[7],它可用于自然或人工生态系统稳定状况的评估[9-10]及环境的监测[11]。有报道称,土壤中的蜘蛛还能参与土壤的修复[12]。通过对江西南昌梅岭景区蜘蛛多样性及形成机制进行调查与分析,期待掌握该景区蜘蛛资源分布的本底情况及区系特点,了解其蜘蛛物种、群落结构与捕食功能多样性规律及其相互间的联动关系,探明其区域特色种的多样性形成原因,最终为该区生物多样性的保护与利用提供理论依据。
1 材料与方法
1.1 研究区概况
该研究集中于江西南昌市梅岭景区的梅岭主峰(A)、洪崖丹井(B)、市植物园(C)、狮子峰(D)、国家森林公园(E)和方志敏陵园(F)6个不同景点。
1.2 研究方法
1.2.1 野外研究方法 通过在2014年相同时段的野外调查、标本采集(定点和随机法、诱捕和人工法相结合)、体积分数为75%酒精浸泡保存,从梅岭主峰、洪崖丹井、市植物园、狮子峰、国家森林公园和方志敏陵园6个不同景点的主要生态类型中获取土壤、地表、树表、岩表、水表等样本,经分离提取,共获得蜘蛛标本784份。标本采集情况详见表1。
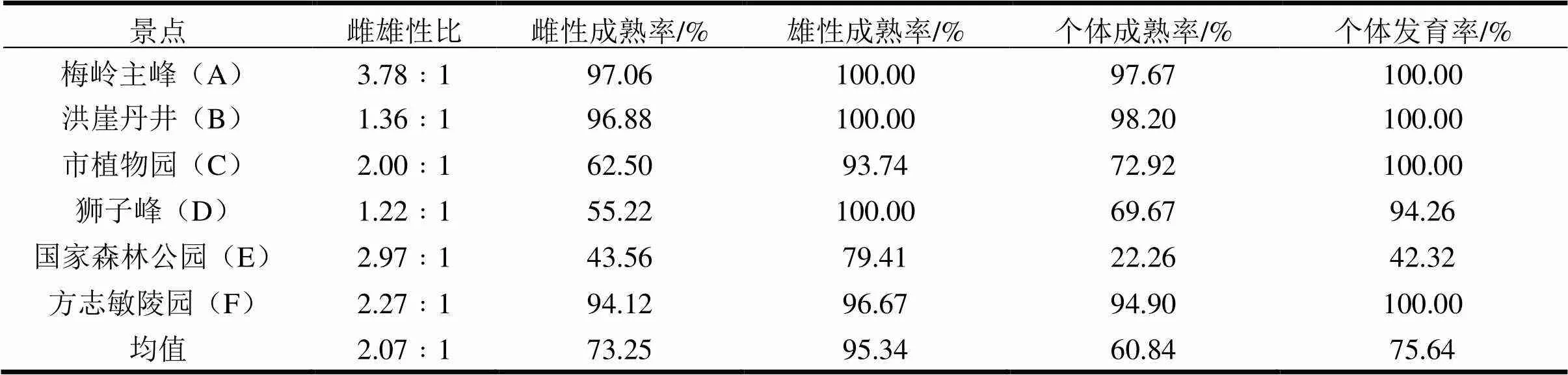
表1 江西南昌梅岭景区6个不同景点蜘蛛标本采集情况
1.2.2 室内研究方法 依据尹长民等(2012)的《湖南动物志,蜘蛛类》等文献资料[4,14-17],在连续变倍体视镜下对蜘蛛眼、纺器、外雌器或触肢器等形态进行识别分类,物种鉴定到种,将雌雄性成蛛(性腺完全成熟)、雌雄性亚成蛛(未完全成熟但性器外观可辨识)和若蛛(形态或出处可辨识)分开计数,在本文分析过程中全部统计。
1.2.3数据处理与分析 本研究涉及的数据处理与分析有:(1)多样性指数:多样性指数是一种反映群落中物种丰富度和个体均匀分布程度的指标[18-19]。(2)聚类分析:聚类分析是从一批样本中的多个观测指标中计算出样本之间的相似程度或距离[20]。(3)主成分分析:主成分分析也称主分量分析,旨在利用降维的思想,把多指标转化为少数几个综合指标(即主成分)[21]。(4)相关性分析:相关性分析是指对两个或多个具备相关性的变量元素进行分析,从而衡量两个变量因素的相关密切程度[21]。本研究涉及的多样性指数的测定和群落间聚类分析采用BioDiversity Pro.软件进行数据处理与分析,而涉及的将各种多样性指数作为变量进行的主成分分析、相关性分析及聚类分析则采用SPSS 17.0软件进行数据处理与分析。
2 结果与分析
2.1 蜘蛛资源名录及区系分布
经鉴定,江西南昌梅岭景区6个不同景点共有蜘蛛27科91属139种,其中有江西新记录种17种,未确定种39种,1个雄性新发现。具体名录及分布详见表2。

表2 江西南昌梅岭景区蜘蛛资源及区系分布
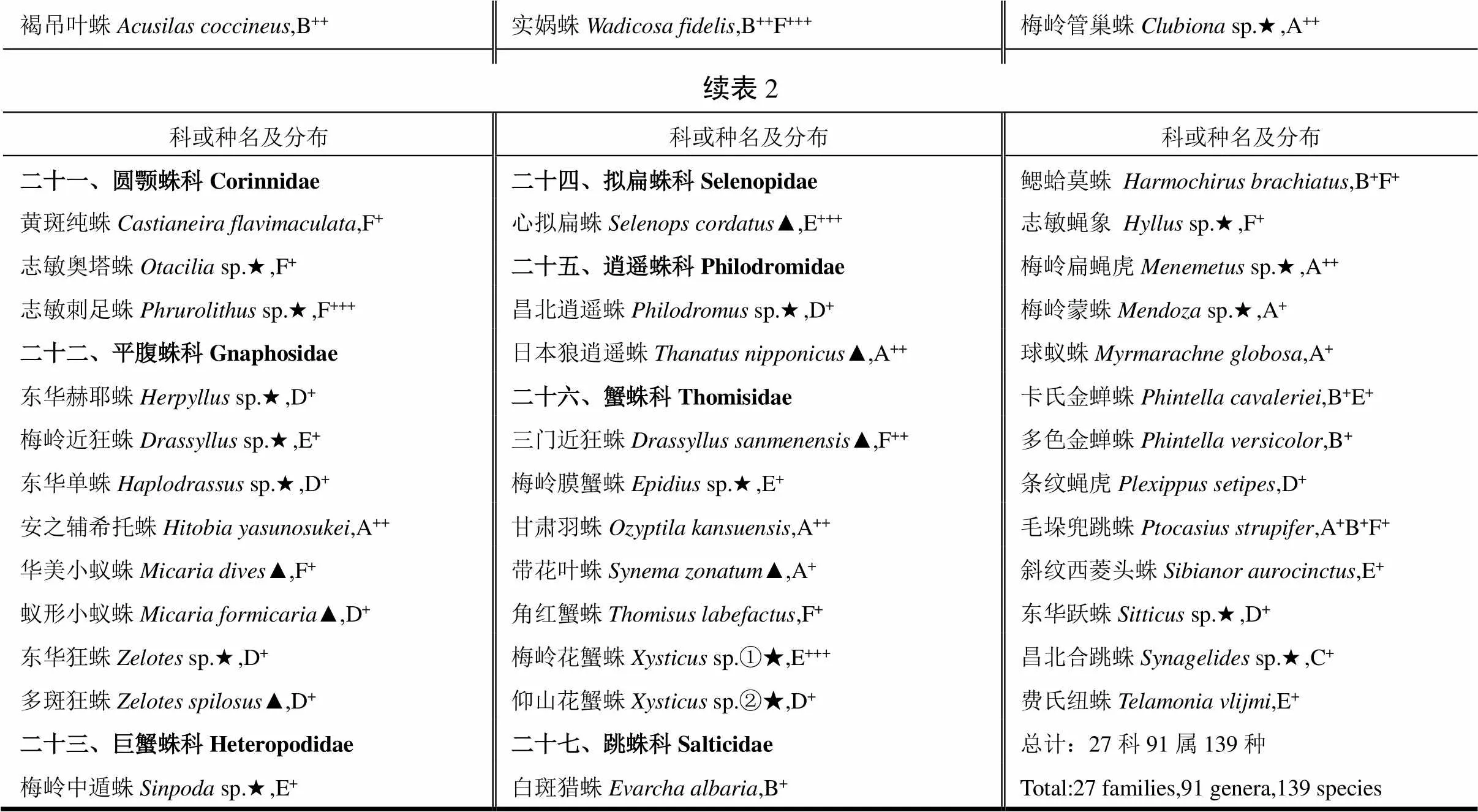
褐吊叶蛛Acusilas coccineus,B++实娲蛛Wadicosa fidelis,B++F+++梅岭管巢蛛Clubiona sp.★,A++ 续表2 科或种名及分布科或种名及分布科或种名及分布 二十一、圆颚蛛科Corinnidae二十四、拟扁蛛科Selenopidae鳃蛤莫蛛Harmochirus brachiatus,B+F+ 黄斑纯蛛Castianeira flavimaculata,F+心拟扁蛛Selenops cordatus▲,E+++志敏蝇象Hyllus sp.★,F+ 志敏奥塔蛛Otacilia sp.★,F+二十五、逍遥蛛科Philodromidae梅岭扁蝇虎Menemetus sp.★,A++ 志敏刺足蛛Phrurolithus sp.★,F+++昌北逍遥蛛Philodromus sp.★,D+梅岭蒙蛛Mendoza sp.★,A+ 二十二、平腹蛛科Gnaphosidae日本狼逍遥蛛Thanatus nipponicus▲,A++球蚁蛛Myrmarachne globosa,A+ 东华赫耶蛛Herpyllus sp.★,D+二十六、蟹蛛科Thomisidae卡氏金蝉蛛Phintella cavaleriei,B+E+ 梅岭近狂蛛Drassyllus sp.★,E+三门近狂蛛Drassyllus sanmenensis▲,F++多色金蝉蛛Phintella versicolor,B+ 东华单蛛Haplodrassus sp.★,D+梅岭膜蟹蛛Epidius sp.★,E+条纹蝇虎Plexippus setipes,D+ 安之辅希托蛛Hitobia yasunosukei,A++甘肃羽蛛Ozyptila kansuensis,A++毛垛兜跳蛛Ptocasius strupifer,A+B+F+ 华美小蚁蛛Micaria dives▲,F+带花叶蛛Synema zonatum▲,A+斜纹西菱头蛛Sibianor aurocinctus,E+ 蚁形小蚁蛛Micaria formicaria▲,D+角红蟹蛛Thomisus labefactus,F+东华跃蛛Sitticus sp.★,D+ 东华狂蛛Zelotes sp.★,D+梅岭花蟹蛛Xysticus sp.①★,E+++昌北合跳蛛Synagelides sp.★,C+ 多斑狂蛛Zelotesspilosus▲,D+仰山花蟹蛛Xysticus sp.②★,D+费氏纽蛛Telamonia vlijmi,E+ 二十三、巨蟹蛛科Heteropodidae二十七、跳蛛科Salticidae总计:27科91属139种 梅岭中遁蛛Sinpoda sp.★,E+白斑猎蛛Evarcha albaria,B+Total:27 families,91 genera,139 species

2.2 多样性作用因子的统计分析
将江西南昌梅岭景区6个不同景点蜘蛛资源名录中的各区域特色种(未确定种,参见表3中标有星号的物种)及其所属科、景点、群落结构类型、捕食功能类型分别列出[13],然后再将表4、表6、表7和表8[13]中它们各自对应的科级物种丰富度指数,所属景点物种丰富度指数、优势度指数、多样性指数和均匀度指数,所属群落结构丰富度指数、优势度指数、多样性指数和均匀度指数,及所属捕食功能类型丰富度指数、优势度指数、多样性指数和均匀度指数共13项参数作为变量,采用SPSS16.0软件对上述各参数数据进行处理,分别做主成分分析、相关性分析及聚类分析。
2.2.1 多样性作用因子的主成分分析 各指数变量的主成分分析表明,上述13个因子对南昌梅岭景区中的区域特色种形成具体作用大小依次为:景点物种均匀度、景点物种丰富度、景点物种优势度、捕食功能均匀度、群落结构丰富度、捕食功能丰富度、群落结构均匀度、群落结构优势度、景点物种多样性、群落结构多样性、捕食功能优势度、捕食功能多样性、科级物种丰富度;前6个为主成分,即景点物种均匀度、景点物种丰富度、景点物种优势度、捕食功能均匀度、群落结构丰富度、捕食功能丰富度是该景区区域特色种多样性形成的主要作用因子。
2.2.2 多样性作用因子的相关分析 各指数变量的相关性分析表明,科级物种丰富度与其它指数相关性均不显著;景点物种丰富度与景点物种多样性、景点物种均匀度极显著相关(<0.01),与捕食功能丰富度显著相关(<0.05);景点物种优势度与景点物种多样性、景点物种均匀度极显著相关(<0.01);景点物种多样性与景点物种多样性、景点物种优势度极显著相关(<0.01);景点物种均匀度与景点物种多样性、景点物种优势度极显著相关(<0.01);群落结构丰富度与群落结构均匀度极显著相关(<0.01);群落结构优势度与群落结构多样性、捕食功能多样性极显著相关(<0.01),与捕食功能优势度显著相关(<0.05);群落结构多样性与群落结构优势度、捕食功能优势度、捕食功能多样性极显著相关(<0.01);群落结构均匀度与群落结构丰富度极显著相关(<0.01);捕食功能丰富度与捕食功能优势度、捕食功能均匀度极显著相关(<0.01),与景点物种丰富度显著相关(<0.05);捕食功能优势度与群落结构多样性、捕食功能丰富度、捕食功能优势度、捕食功能均匀度极显著相关(<0.01),与群落结构优势度显著相关(<0.05);捕食功能多样性与群落结构优势度、群落结构多样性极显著相关(<0.01);捕食功能均匀度与捕食功能丰富度、捕食功能优势度极显著相关(<0.01)。故此,除科级物种丰富度与其它指数相关性均不显著外,各类多样性指数变量间多为极显著相关(<0.01)或显著相关(<0.05)。
2.2.3 多样性作用因子的聚类分析 各指数变量的聚类分析表明,昌北毛丛蛛(sp.)与东华指蛛(sp.)、梅岭花蟹蛛(sp.)与昌北舞蛛(sp.)、梅岭中遁蛛(sp)与昌北艾瓦蛛(sp.)、梅岭管巢蛛(sp.)与梅岭隙蛛(sp.)、昌北旋隙蛛(sp.)与昌北毛丛蛛(sp.)等之间的距离较近,因为它们亲缘关系很远,隶属于不同科属,所以其演化过程中生态环境等外在因素就成为它们趋近的主要动力;而梅岭膜蟹蛛(sp.)与梅岭花蟹蛛(sp.)、东华单蛛(sp.)与东华狂蛛(sp.)、东华赫耶蛛(sp.)与东华单蛛(sp.)、志敏奥塔蛛(sp.)与志敏刺足蛛(sp.)等之间的距离较远,因为它们亲缘关系较近,隶属于同科或属,故其演化过程中行为遗传方式等内在因素就是它们趋远的主要动力。此外,其它区域特色种形成的动力则来源于内外因(即遗传与环境)的综合作用。
3 讨论与小结
本次对江西南昌梅岭6个景点蜘蛛资源的调查,共采集到标本784份,经分类鉴定,它们分别隶属于27科91属139种,其中有17个江西新记录种、39个未确定种和1个雄性新发现。与周边类似景区的蜘蛛资源相比[22-23],南昌梅岭景区蜘蛛资源较为丰富,其科属种数处于偏高水平。并且,该景区分布较广的蜘蛛以地穴型和地面游猎型种类为主,如触形大疣蛛()沟渠豹蛛()、雾豹蛛()、细毛水狼蛛()、田野安蛛()、草栖毛丛蛛()等,常见的结网型蜘蛛只有棒络新妇()等极少种类,另外,区域特色种各科均有分布。这与该山域绵延开阔(位于赣中鄱阳湖冲积平原且周边无障碍)、东北向鄱湖水气条件充分(临近鄱阳湖西南角)、温湿度适宜(水气交流频繁且四季分明)、植被类型多样(灌丛草甸为主)、昆虫相复杂(土壤和地面昆虫丰度较高)及人为干扰频繁等密切相关。
因为蜘蛛与植物、昆虫关系密切,它既与植物存在特定的群落结构,也与昆虫间存在紧密的捕食关系。本文依据蜘蛛栖息与捕食等生态行为特点对其群落结构类型进行区分,旨在更好地反映其对时空景观格局变化的适应[13,24-26]。同时,根据Pekár等[27]对蜘蛛捕食昆虫的统计与食性的演化分析及作者野外调查的结果[5]将蜘蛛按捕食功能区分为广食性、狭食性和寡食性等类型[13,24-26],则突显了蜘蛛与昆虫协同进化[29]的不争事实。虽然狭食性区分的依据是其偏好猎物而不是潜在猎物,并且猎物多样性和丰度较高时蜘蛛狭食性演化情形明显[5,27-28],但特定情形下狭食性蜘蛛也会略为改变其食性,以适应新环境。换言之,对蜘蛛群落结构和捕食功能多样性的分析有助于该物种区系分布和多样性形成机制等问题的分析和理解。
南昌梅岭景区蜘蛛多样性作用因子的主成分分析表明,景点物种均匀度、景点物种丰富度、景点物种优势度、捕食功能均匀度、群落结构丰富度、捕食功能丰富度是其区域特色种多样性形成的6个主要作用因子。而多样性作用因子的相关性分析表明,各因子间普遍相关;多样性作用因子的聚类分析表明,区域特色种形成的动力来源于内外因(即遗传与环境)的综合作用。这些分析结果表明,在对该景区蜘蛛多样性形成原因与方式的研究中,各种指数的选用是合适且有意义的,这些多样性形成的主要因素与国外相关研究[30-35]的结果相一致。
[1] 王洪全, 颜亨梅, 杨海明.中国稻田蜘蛛群落结构研究初报[J]. 蛛形学报, 1999(8): 95-105.
[2] 尹长民. 中国蜘蛛生态学研究概况[J]. 蛛形学报, 1999(8): 122-127.
[3] Platnick I N. The world spider catalog[EB/OL]. (2000-01-15) http://www.wsc.nmbe.ch[2019-10-20].
[4] 李枢强, 林玉成.中国生物物种名录, 蜘蛛目[M]. 北京: 科学出版社, 2016: 1-549.
[5] 陈连水, 袁凤辉, 周谷春, 等. 江西省蜘蛛资源的最新研究报告[C]. 南昌: 第十届南方六省(湘鄂赣粤桂琼)动物学学术研讨会论文摘要, 2018: 37.
[6] 申效诚, 张保石, 张锋, 等. 世界蜘蛛的分布格局及其多元相似性聚类分析[J]. 生态学报, 2013(33): 6795-6802.
[7] Wolfgang N. Spider ecophysiology[M]. Berlin: Springer-Verlag Berlin and Heidelberg GmbH & Co. K, 2013:2-8.
[8] 谢国文, 颜亨梅, 张文辉.生物多样性保护与利用[M]. 长沙: 湖南科学技术出版社, 2000: 36.
[9] Patrick M, Alain C, Frédéric Y. Spiders (Araneae) useful for pest limitation and bioindication[J]. Agriculture Ecosystem & Environment, 1999(74): 229-273.
[10] Maija Š, Voldemārs S. Diversity of grass-dwelling spiders (Arachnida: Araneae) in calcareous fens of the Coastal Lowland, Latvia[J]. Journal of Insect Conservation, 2014(18): 757-769.
[11] Xiao-li S, Yu P, Hose G C, et al. Spider Webs as Indicators of Heavy Metal Pollution in Air[J]. Bulletin of Environmental Contamination and Toxicology, 2006(76): 271-277.
[12]曹阳, 高梅香, 张雪萍, 等. 黑龙江省不同纬度梯度农田大型土壤动物群落分布特征[J]. 生态学报, 2017(37): 1677-1687.
[13] 袁凤辉, 陈连水, 周谷春, 等. 江西南昌梅岭景区蜘蛛多样性的比较研究[J]. 生物灾害科学, 2017(40): 143-150.
[14] 尹长民, 彭贤锦, 颜亨梅, 等. 湖南动物志, 蜘蛛类 (上下册) [M]. 长沙: 湖南科学技术出版社, 2012: 1-1590.
[15] Song D X, Zhu M S, Chen J. The spiders of China[M]. Hebei: Science and Technology Publishing House, 2000: 1-290.
[16] 朱明生, 王新平, 张志升. 中国动物志, 蛛形纲, 蜘蛛目, 漏斗蛛科, 暗蛛科[M]. 北京: 科学出版社, 2017: 1-727.
[17] 张志升, 王露雨. 中国蜘蛛生态大图鉴[M]. 重庆: 重庆大学出版社, 2017: 1-938.
[18] 马克平. 生物群落多样性的测度方法.生物多样性研究的原理与方法[M]. 北京: 中国科学技术出版社, 1994: 66-68.
[19] Anne E M. 生物多样性测度[M]. 张峰, 译.北京: 科学出版社, 2011: 67-86.
[20] 赵志模, 郭依泉. 群落生态学原理与方法[M]. 重庆: 科学技术文献出版社, 1989: 77-89.
[21] 李春喜, 姜丽娜, 邵云, 等.生物统计学(第五版) [M]. 北京: 科学出版社, 2016: 112-220.
[22] 陈连水, 黄芳, 袁凤辉, 等. 江西庐山景区蜘蛛资源的调查初报[J]. 生物灾害科学, 2016, 39(3): 149-152.
[23] 袁凤辉, 陈连水, 饶军, 等. 福建武夷山景区蜘蛛资源的初步调查[J]. 蛛形学报, 2016, 25(1): 47-49.
[24] 陈连水, 袁凤辉, 周谷春, 等.江西相山铀矿蜘蛛群落结构与捕食功能多样性研究[J/OL]. [2020-02-03]. http://www.docin. cont/p-1151138047.html.
[25] 黄珍, 陈连水, 吴心怡, 等.江西境内触形大疣蛛(蜘蛛目:异纺蛛科)种系形态的区系演化[M/CD]. 中国科技论文在线精品论文, 2015(8): 2067-2074.
[26] 陈连水, 胡丽萍, 袁凤辉, 等. 触形大疣蛛区域性演化模式的探讨[J]. 江西科学, 2019, 37(3): 390-396.
[27] Stano P, Jonathan A C, Todd A. Blackledge,evolution of stenophagy in spiders (Araneae): evidence based on the comparative analysis of spider diet[J]. Evolution, 2012(66): 776-806.
[28] Stano P, Søren T. Trophic specialisation in a predatory group: the case of prey-specialised spiders (Araneae)[J]. Biological Reviews, 2015(90): 744 -761.
[29] Haynes K F, YearganK V, GemenoC. Detection of prey by a spider that aggressively mimics pheromone blends[J]. Journal of Insect Behavior, 2001(14): 535-544.
[30] Matthew H G. Determinants of web spider species diversity: vegetation structural diversity vs. prey availability[J]. Oecologia, 1984(62): 299-304.
[31] Alberto C, David H W. Influence of fine woody debris on spider diversity and community structure in forest leaf litter[J]. Biodiversity and Conservation, 2009(18): 3705-3731.
[32] Huang P S, Lin H C, LinC P, et al. The effect of thinning on ground spider diversity and microenvironmental factors of a subtropical spruce plantation forest in East Asia[J]. European Journal of Forest Research, 2014(133): 919-930.
[33] Roland H, Tibor M, Csaba S, et al. In stable, unmanaged grasslands local factors are more important than landscape-level factors in shaping spider assemblages[J].Agriculture, Ecosystems & Environment, 2015(208): 106-113.
[34] Roland H, Szabolcs L, Csaba S, et al. The effect of prey availability on spider assemblages on European black pine () bark: spatial patterns and guild structure[J]. Canadian Journal of Zoology, 2005(83): 324-335.
[35] Andreas S, Helge B, Walter D, et al. Tree diversity promotes functional dissimilarity and maintains functional richness despite species loss in predator assemblages[J].Oecologia, 2014(174): 533-543.
Investigation and Its Formation Cause Analyse of the Spider Diversity in the Meiling Scenic Area of Nanchang, Jiangxi
CHEN Lian-shui1, YUAN Feng-hui1, ZHOU Gu-chun2, RAO Jun1, DU Shi-sheng1, ZHANG Yan-yan1
(1. Department of Biology,East China University of Technology, Nanchang 330013, China; 2. College of Life Science, Gannan Normal University,Ganzhou 341000, China)
In this paper, the diversity of spiders in 6 typical scenic spots in the Meiling Scenic area of Nanchang City, Jiangxi Province was investigated, and the detailed distribution list of spider resources was identified and obtained. Combined with the results of comparative study on its spider diversity that had been reported on the other magazine, the formation cause and mode of the 39 undetermined species on list were revealed as a regional characteristic species by using a total of 13 indexes of their species richness, species diversity, community structure diversity and predation functional diversity as variables to analyze the main components, correlation and clustering of action factors. The results showed that the species evenness, the scenic spot species richness, the scenic spot species dominance, the predation functional evenness, the community structure richness and the predation function richness were the six main factors for the formation of regional characteristic species, and the various factors were common correlation,then the ecological and genetic factors each focused on their formation.
Nanchang; the Meiling Scenic Area; spider; diversity; formation cause.
Q958
A
2095-3704(2020)01-000048-06
2020-02-27
国家自然科学基金项目(31201718)、江西省自然科学基金项目(20142BAB204011)和国家级大学生创新创业训练计划项目(2017JGSH40-21)
陈连水(1965—),男,副教授,硕士,主要从事蛛形及生态学研究,dhlianshui81@163.com。
陈连水, 袁凤辉, 周谷春, 等. 江西南昌梅岭景区蜘蛛多样性调查及成因分析[J]. 生物灾害科学, 2020, 43(1): 48-53.
10.3969/j.issn.2095-3704.2020.01.10
