不同种源杉木树轮α纤维素δ13C对年气候因子的响应
2020-04-21刘小英段爱国张建国张雄清朱安明
刘小英,段爱国,2*,张建国,2,张雄清,朱安明
(1. 中国林业科学研究院林业研究所,国家林业和草原局林木培育重点实验室,北京 100091;2. 南京林业大学南方现代林业协同创新中心,江苏 南京 210000;3. 中国林业科学研究院林业科技信息研究所,北京 100091)
造林树种种源选择已成为人类应对气候的重要途径,探讨不同地理种源主要造林树种树轮指标对气候变化的响应及评价未来气候变化对林木种源生长的影响已成为亟待解决的重要科学问题。林木种源是指取得种子或其他繁殖材料的地理来源或原产地[1]。地理种源的变异是树木长期气候环境适应性进化的产物,从遗传角度看,不同地理种源经过自然选择基因型组成会发生变化,形成固有的遗传基础[2-4],因此,在引种栽培时往往采取“就近原则”,选择同一种源或同一种源区的幼苗进行栽植[5]。然而,有研究者通过对不同种源山毛榉(Fagus sylvaticaL.)应对气候变化的响应情况进行研究,发现不同种源山毛榉对异地(栽植地)的气候具有不同的响应特征,而最适宜种源并不满足“就近原则”[6-7]。
树轮稳定碳同位素(δ13C)技术被广泛应用于生理生态、气候变化、水文等研究领域,现已逐步成为了解生物与其生存环境相互关系的工具;而树木是如何应对气候变化依旧被气候学家关注,与树轮宽度、木材密度相似,树轮稳定碳同位素指标对不同尺度上气候变化的响应存在很大的差异[8]。目前,针对气候响应研究的气候因子主要有平均气温[9]、降水[10]、相对湿度[11-12]、最高气温、最低气温[13]、最小相对湿度及日照时数[6]。有研究表明,树轮稳定碳同位素比树轮宽度指标对气候变化的响应更灵敏。在种源层面上,树木种源对环境的影响具有差异性,研究发现,不同种源的白刺(Nitraria tangutorumBobr.)对不同梯度的水分响应存在较大的区别[14];不同种源的落叶松(Larix gmelinii(Rupr.) Kuzen.)对原产地环境条件的响应也出现差异[15]。不同种源树木树轮的早材、晚材、整轮宽度对年平均气温和年最高气温较敏感[16]。杉木(Cunninghamia lancealata(Lamb.)Hook.)是我国重要的乡土针叶用材树种[17],其生长呈现明显的地理纬度变异,针对杉木树轮指标在种源层面上响应气候变化的研究相对较少,而将高灵敏度的树轮稳定碳同位素指标引入不同种源树木生长对气候变化响应的研究国内外未见报道。
鉴于此,本研究选择设置于广西柳州的34年生杉木种源试验林中的30个代表性地理种源,探讨其树轮α纤维素中稳定碳同位素与年际气候因子间的关系,以期筛选出关键气候因子,为未来气候变化条件下种源层面上区域杉木的培育提供科学支撑。
1 研究区自然概况
杉木种源试验林位于广西柳州六峰山林场,由中国林科院林业研究所主持,全国杉木地理种源试验协作组于20世纪70年代末开展的第2次杉木全分布区种源试验时期布设[18]。依据杉木产区的划分,其属于杉木的中心产区。试验林于1981年春季,采用1年实生苗造林。种源试验林所处林地较平坦,随机区组设计。造林行间距2 m×2 m,每个小区4株,重复10次,进行了1次疏伐,至2013年秋季,杉木种源试验林保留密度为1 261株·hm-2,平均胸径20.14 cm,平均树高13.45 m。试验林地处桂中丘陵地区 (23°42′ N,109°50′ E),年平均气温21.1℃,年降水量1 418.5 mm,年蒸发量1 969.1 mm,相对湿度76%,全年日照时数1 823.7 h。
2 研究材料与方法
2.1 树轮资料
结合种源林商业采伐,对所有种源保留单株在20 cm高度处截取了圆盘,考虑地理分布的代表性及圆盘数量大于10个,选取贵州六枝(GZl)、湖南双牌(HNs)以及福建永安(FJy)等30个地理种源各11个单株,合计330个单株进行分析,其中,中心产区10个种源,一般产区5个种源,边缘产区15个种源。平均胸径20.33 cm,平均树高13.57 m。应用LignoStation年轮分析仪对不同种源各单株圆盘进行扫描,分别以树木的南、北2个方向进行测量,应用年轮分析仪自带软件进行年轮界线的划分,并量测树轮自髓心往外各年轮宽度,后借助交叉定年软件COFECHA程序对所有定好年的序列作检验。
2.2 α-纤维素提取
将树轮圆盘放在干净的玻璃板上,选择向阳的方位[19],在显微镜下用大头针标注经交叉定年的树轮。考虑到δ13C值会受到幼龄效应的影响[20],在取样时去除前6年的树轮。利用干净的刻刀进行树轮的逐年剥离,将同一种源的11个圆盘同一年的年轮样品剥取后混合在一起,放入高速离心球磨仪将样品粉碎至过60目筛(300 μm)。按照有机溶剂(苯:无水乙醇=2∶1,丙酮180 mL)萃取、漂白(次氯酸钠∶乙酸=3∶2)及碱化(10%与17%的氢氧化钠依次处理)等步骤提取α-纤维素[21-22]。用真空冷冻干燥机(-45℃)进行冻干处理,采用快速核酸提取仪进行α-纤维素粉末的混合。
2.3 碳稳定同位素(δ13C)的测量
采用仪器Thermo Finnigan DELTA plus XP气象同位素质谱仪测量杉木全木以及α-纤维素的δ13C的值,该过程在植被与变化国家重点实验室完成。用百万分之一天平(Starorius)称量粉末α-纤维素(0.20±0.05)mg,用锡囊将样品包裹,卷好密封后上机测试,得到13C/12C值(相对VPDB标准),即δ13C。整个碳同位素(δ13C)测量精度≤0.15‰。

式中:Rsample表示样品中的重轻同位素峰度之比,Rstandard是国际通用标准的重轻同位素峰度之比[23]。
2.4 气候数据来源

图 1 广西来宾气象站月平均温度与总降水量变化(1985-2013)Fig. 1 The change trend of the mean temperature and year total precipitation of each month of the climate station of Laibin, Guangxi (1985-2013)
本实验采用广西来宾气象站点的数据(图1),该站点距离种源试验林场100 km,中间无高山阻隔。所用气象资料包括1985—2013年的年平均气温(T)、年最高气温(Ta)、年最低气温(Ti)、年降水量(P),年平均相对湿度(RH)、年最小相对湿度(RHi)和日照时数(S)(气候资料来源于国家气象信息中心来宾气象站点的数据)。经检验,来宾站点气象数据可靠,无明显突变,该站点气候数据代表了当地气候变化趋势。
3 结果与分析
3.1 不同种源杉木δ13C的统计特征
对30个种源交叉定年后,选用去除大气CO2浓度的年表作为后续分析年表,年表统计特征包括各种源树轮δ13C的最小值、最大值、平均值、标准差、一阶自相关系数、二阶自相关系数及平均敏感度。
从杉木树轮α-纤维素δ13C的测定结果可知:1987—2013年杉木树轮δ13C的变幅为-20.997‰~-25.910‰(表1)。δ13C序列随时间呈波动变化,所有种源的δ13C平均值达-24.022‰,标准差变幅为0.24‰~0.47‰。总体上,不同种源树轮样品δ13C均值比较离散,为基于树轮稳定碳同位素探讨不同种源对气候变化的响应提供了必要条件。
通过对不同地理种源杉木年轮 α-纤维素δ13C序列进行自相关分析(表1)发现:不同地理种源杉木的一阶、二阶自相关系数较大,一阶自相关系数为 0.181~0.866,而二阶自相关系数为0.044~0.749,且大部分种源的一阶和二阶自相关系数超过0.4。由此可以说明,大部分种源杉木树轮α-纤维素中δ13C值在响应气候变化时存在明显“滞后效应”,故选择当年、前1年、前2年的气候因子进行相关分析。
3.2 不同种源杉木树轮δ13C值与气候因子相关性分析
3.2.1 杉木树轮δ13C与年气象因子的相关分析从图2可看出:大部分杉木种源树轮α-纤维素中δ13C与年平均气温、年最高气温、年最低气温呈显著正相关,与年平均相对湿度、年最小相对湿度呈显著负相关,而与年降水、日照时数及湿润度指数不显著相关。这是因为树轮α纤维素中δ13C的累积主要通过植物的光合作用进行,而温度的升高,有利于光合作用过程中酶活性的提高,能更有效的吸收大气中的CO2,有效促进光合作用,从而使树轮稳定碳同位素可以更灵敏的响应气候的变化。杉木生长需要的湿度相对较大,当植物叶片处于干燥的环境时,水分的减少导致气孔导度降低,限制了叶片中CO2的供给,从而树轮α纤维素中δ13C增大,致使树轮稳定碳同位素可以更灵敏的响应气候的变化。
不同产区杉木树轮的δ13C对气候因子的响应灵敏程度存在差异。图2表明:对于湖南安化、福建南平、福建崇安、广西三江、广东乐昌、广西融水、湖南祁阳、江西遂川、湖南双牌、广东始兴等杉木种源而言,不同种源树轮的δ13C与当年的最小相对湿度和前2年的平均温度、最低气温、最高气温及平均相对湿度的相关系数较高,其中,与前2年的平均相对湿度呈显著负相关(p<0.05),与当年的最小相对湿度呈极显著负相关(p<0.01)。对于四川汶川、浙江龙泉、江西修水、四川古蔺、云南西畴等杉木种源而言,不同种源树轮δ13C与当年的最低气温、前1年的最小相对湿度、平均气温及前2年的平均相对湿度、最高气温的相关系数较高,其中,与前1年最小相对湿度呈极显著负相关(p<0.01),与当年的平均相对湿度呈显著负相关(p<0.05);而广东紫金、贵州六枝、湖南新宁、江苏句容、广西博白、福建永安、湖北谷城、陕西南郑、云南屏边、湖北罗田、河南商城、安徽东至、贵州麻江、浙江丽水、福建武平的杉木种源,不同种源树轮δ13C值与当年的最小相对湿度、平均相对湿度和前2年的平均气温、最低气温、最高气温的相关系数较大,其中,与当年的最小相对湿度和平均相对湿度呈极显著负相关(p<0.01),且与当年的最小相对湿度相关系数较大。不同产区原产地的气候、地貌、土壤等立地条件均不相同,由于受到不同因素的影响,不同杉木种源的遗传基础出现差异,当将其引种到同一气候条件下进行栽培时,这种差异性导致了不同种源对栽植地气候因子的差异性响应。

表 1 1987-2013年不同种源杉木树轮稳定碳同位素(δ13C)序列的基本统计特征Table 1 Basic statistical characteristics of stable carbon isotope sequences of different tree provenances in China from 1987 to 2013
不同地理种源杉木树轮的δ13C与当年降水呈不显著相关(图2)。种源FJw与当年最低气温呈现出负相关性,而大多数种源与当年、前1年及前2年最低气温呈显著或极显著正相关,相关系数最大达0.677(p<0.01)。对最小相对湿度的相关性分析中发现,除种源Ync相关性不显著外,其余种源均与当年、前1年及前2年最小相对湿度呈显著或极显著负相关,相关系数最大达-0.744(p<0.01)。与日照时数的相关性分析中发现,仅少数种源与当年、前1年及前2年呈显著正相关,相关系数最大达0.457(p<0.01)。不同地理种源杉木树轮δ13C与平均气温的相关性分析中发现(图2),大部分种源与当年、前1年及前2年呈显著或极显著正相关,而种源GXb与平均气温呈显著负相关,相关系数可达-0.418(p<0.05)。多数种源与当年、前1年及前2年平均相对湿度呈显著或极显著负相关,而种源GDz、GXs及Ynb与前1年呈显著或极显著正相关。不同地理种源杉木树轮δ13C与前2年最高气温的相关性分析中发现(图2),种源GXs与其呈现极显著正相关,相关系 数 0.569(p<0.01) , 种 源 SXn、 FJw、 HBt、HNq、HNs、GDs、GZm、Ynb、SCc与最高气温呈现出显著的正相关关系,相关系数可达0.447(p<0.05)。
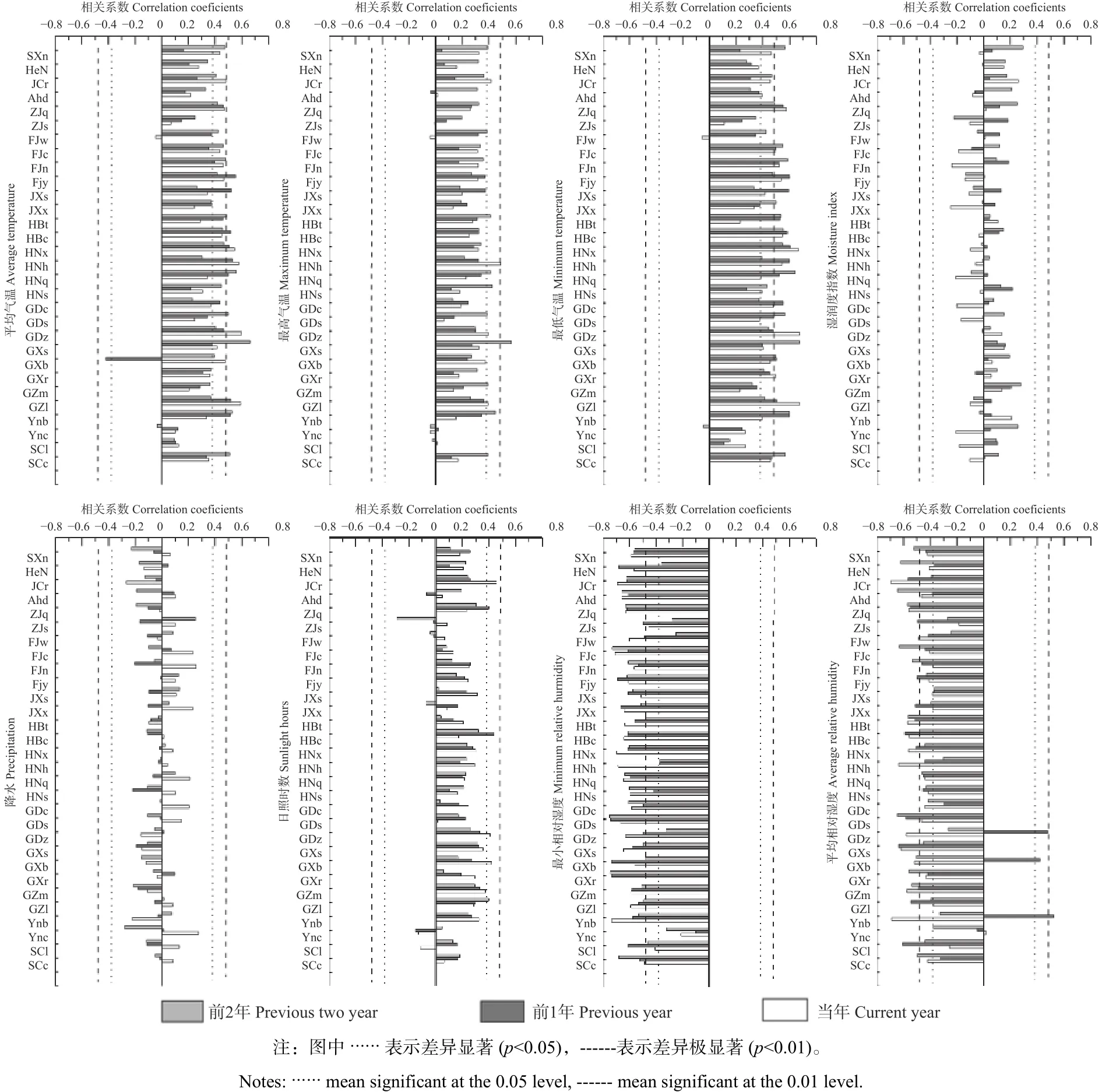
图 2 杉木树轮δ13C与年际气候因子相关分析Fig. 2 Correlation analysis between δ13C and annual climatic factors of fir tree rings
3.2.2 不同地理种源杉木树轮δ13C与当年气候因子相关性随经纬度的变化 从杉木地理种源树轮δ13C与当年气候因子相关性随经度的变化趋势(图3)可知:杉木种源树轮δ13C与年降水的相关性受经度影响较小(r=0.028),与年平均气温、最低气温、最高气温、日照时数的相关性随经度的增加总体上呈先增强后减弱的变化规律,且在110~115°E间正相关性最强。杉木种源树轮δ13C与平均相对湿度及最小相对湿度的负相关性随经度的增大总体呈先增强后减弱的趋势,且在110~115° E间呈现最强负相关性,δ13C与最小相对湿度的负相关性随经度的这种先增强后减弱的变动趋势可由二次多项式描述,且达显著水平(r=-0.462)。从稳定碳同位素与各年际气候因子的相关系数随纬度的变化趋势图(图3)可以发现,杉木种源树轮δ13C与年降水量的相关系数受种源所处纬度的影响较大,随纬度的增加呈先上升后下降的显著变化趋势(r=0.434),而不同种源树轮δ13C与年平均气温、最低气温、最高气温、平均相对湿度、日照时数、最小相对湿度的相关关系则受纬度影响较小。
3.3 不同种源杉木树轮α-纤维素中δ13C与气候因子的多元回归分析

图 3 δ13C与当年各年际气候因子的相关系数随着经(纬)度的变化散点图Fig. 3 Scatter diagram of correlation coefficients between the δ13C value and inter-annual climate factors along with longitude and latitude of provenances
为了更深入地了解气候因子对不同种源杉木树轮δ13C的影响情况,以树轮α纤维素δ13C为因变量,自变量选取相关性较强的当年最小相对湿度(X1)及前2年的平均气温(X2)、最低气温(X3)、最高气温(X4)和平均相对湿度(X5),回归方法选择逐步回归法,置信度<0.05,输出结果见表2。
从表中2中α-纤维素δ13C序列的逐步回归分析可知:在剔除了对α-纤维素中δ13C序列影响不显著的因子后,HNh、FJc、GXs、GDc、GXr、HNq、JXs、HNs、GDs、ZJq、JXx、GDz、GZl、HNx、JCr、FJy、HBc、SXn、Ynb、HBt、HeN、Ahd、GZm、ZJs、FJw等25个种源树轮α-纤维素δ13C序列受气候因子当年最小相对湿度影响显著,其中,部分种源同时受到前2年平均相对湿度或前2年最低气温的显著影响,R2处于0.176~0.627之间,说明以当年最小相对湿度为主的气候因子对杉木不同地理种源树轮α-纤维素中δ13C序列的贡献量为17.6%~62.7%;而福建南平(FJn)和四川汶川(SCc)种源均分别与前2年的最低气温和平均相对湿度呈显著正线性及负线性相关,广西博白(GXb)种源与前2年平均气温呈显著负线性相关,云南西畴(Ync)、四川古蔺(SCl)种源与前2年平均相对湿度具显著负线性相关关系。

表 2 δ13C与年值气候因子多元回归分析模型Table 2 Multiple regression analysis model between δ13C values and yearly climate factors
4 讨论
不同种源杉木(1987—2013)树轮α纤维素中稳定碳同位素(δ13C)的变幅为-20.997‰~-25.910‰,且表现出较强的一阶自相关,相关系数为0.181~0.866,表明广西柳州不同种源杉木树轮δ13C的年际变化相对稳定,能较好的呈现环境的变化,这与靳翔等[9]对岷江冷杉树轮δ13C变幅及年际变化的研究结果相一致。
本研究发现,大部分杉木种源δ13C与年平均气温、最高气温、最低气温呈显著正相关,与最小相对湿度、平均相对湿度呈极显著负相关,而与年日照总时数呈不显著正相关。杉木不同种源树轮δ13C与气候因子的这种正负相关性与对亚热带树种猴面包树[24]及亚高山暗针叶林[25]的研究相似。
研究还发现,不同种源的杉木对气候因子响应的灵敏度不同,可见,影响树轮中δ13C值的因素并不单一,是多个因子的综合效应[26]。树轮α纤维素中δ13C值与叶片气孔导度及环境温度相关[13],而叶片是通过感知环境中水分多少来调节气孔导度,当叶片处于相对干燥的环境中,水分的减少导致气孔导度降低,限制了叶片CO2的供给,最终导致树轮中δ13C值增大[27]。温度是影响CO2分馏的主要条件,当温度低于平均温度,且二者之间差值较大时,将会影响树轮中碳的固定[28]。通过对杉木树轮中δ13C值与当年、前1年、前2年气候因子相关性的分析可以看出,17个种源对前2年平均气温、15个种源与前2年最低气温、21个种源与前2年最高气温以及11个种源与前2年的平均相对湿度相关系数较大,而14个种源与当年最小相对湿度呈显著负相关性,除了广东紫金、贵州六枝和湖南新宁的3个种源树轮δ13C值对气候因子的响应是即时效应外,其余的种源对气候因子的响应均存滞后效应。由此可以看出,各气候因子对杉木树轮中稳定碳同位素的影响是长期积累的结果,这与在低海拔地区树轮稳定碳同位素对年气候因子的响应出现明显滞后效应的结论一致[29]。
通过对相关系数较高的气候因子进行逐步回归分析,发现除了福建南平(FJn)种源与前2年的最低气温和平均相对湿度呈线性关系,广西博白(GXb)种源与前2年平均气温呈线性关系,云南西畴(Ync)、四川古蔺(SCl)种源与前2年平均相对湿度呈线性关系外,其余26个种源杉木树轮α纤维素中δ13C值随当年最小相对湿度的增大而出现减小的趋势。因此,当年最小相对湿度可以作为影响杉木树轮α纤维素中δ13C的主导气候因子。在湿度较大的环境中,杉木往往生长较好,其有机物的分馏作用越强,导致树轮中δ13C减小[28]。
5 结论
中亚热带柳州地区不同地理种源杉木树轮α-纤维素中δ13C对同一气候因子的响应方向基本一致,大部分种源树轮δ13C与平均相对湿度和最小相对湿度呈负相关,与平均气温、最高气温和最低气温呈显著正相关,与日照时数、降水和湿润度指数的相关性不大,且有90%的杉木种源树轮δ13C对气候的响应存在明显滞后效应。不同地理种源杉木树轮α-纤维素中δ13C对最小相对湿度这一气候因子的响应较灵敏,从生物学基础上阐明了树轮稳定碳同位素的固定、分馏受到生长环境中最小相对湿度的影响较大,而对年降水量多少的依赖性较小。研究结果有效的填补了种源层面上树轮稳定碳同位素对气候响应研究的不足,也为未来气候变化条件下杉木人工林健康培育提供了依据。
