硬头黄竹花的形态与结构研究
2020-04-16王一方初彩华张月高王曙光
李 娟,王一方,初彩华,詹 卉,张月高,王曙光*
(1. 西南林业大学 云南省高校林木生物技术重点实验室 云南 昆明 650224;2. 西南林业大学 生命科学学院 云南 昆明 650224)
竹类植物的繁殖方式多数以无性繁殖为主,在自然条件下大部分竹种需经历120年或更长的时间才能开花,因而竹类植物的开花现象也极少[1],这导致竹类植物通过生殖器官进行形态学、解剖学、系统分类等方面的研究皆遇到极大的阻碍,尤其是在胚胎学方面的研究更加困难。目前,国内外有关竹类植物花的解剖学已经有少量研究,但仅对少数竹种的生殖生物学进行过详细描述,如孝顺竹(Bambusa multiplex (Lour.) Raeusch. ex Schult.)、绵竹(Bambusa intermedia Hsueh et Yi)、青丝黄竹(Bambusa eutuldoides McClure var. viridi-vittata (W.T. Lin) Chia)、异叶苦竹(Pieioblastus simonii f. heteophyllus Muroi)、绿竹(Dendrocalamopsis oldhami(Munro)Keng)、毛竹(Phyllostachys heterocycla(Carr.) Mitford cv. Pubescens)、鹅毛竹(Shibataea chinensis Nakai)、巨龙竹(Dendrocalamus sinicus Chia et J.L.Sun)、雷竹(Phyllostachys praecox C.D. Chu et C. S. Chao 'Prevernalis')等竹种[2-11]。
硬头黄竹(Bambusa rigida Keng et Keng f.)属禾本科(Gramineae)箣竹属(Bambusa)竹种[12]。主要分布在我国的广东省、贵州省、四川省、江西赣州等地,其秆通直,材质硬且坚厚,可作竹筷、竹凉席等日用品开发,在用材和造纸方面前景广阔[13-14]。以往国内外对硬头黄竹的研究主要集中在形态与理化特性、遗传多样性及解剖结构等方面[15-23],缺乏对其具体花器官及胚胎发育方面的细致研究。本研究通过观察硬头黄竹花的基本结构,对硬头黄竹的分类特征描述进行补充,并对硬头黄竹雌雄配子体发育进行观察,包括子房形态解剖结构以及雌雄配子体发育过程等,总结其胚胎发育规律,分析硬头黄竹结实率较低的原因,为硬头黄竹胚胎学研究积累原始资料,并为硬头黄竹的育种工作打下基础。
1 材料与方法
实验所用材料于2017年4月采自西南林业大学珍稀竹种园。将不同发育阶段的小穗和花枝固定于50%FAA固定液中,进行真空抽气备用。
将固定处理后的小穗使用体视解剖镜(Olympus HO11)进行不同发育时期的结构解剖、测量及拍照。然后再将解剖后的花药和子房,浸蜡包埋后,旋转切片机进行切片,切片厚度为6~8 μm。利用番红-固绿双重染色,加拿大树胶封片[24]。在Nikon-ECLIPSE50显微镜下进行观察和拍照,实验数据使用Excel 软件进行分析。
2 结果与分析
2.1 硬头黄竹花序和小穗的形态结构
硬头黄竹的花序轴是由不同的节和节间营养枝生长发育而来,小穗生于花枝各节,呈淡黄绿色(图1A)。小穗底部具有潜伏芽,柄几无,为假小穗,呈簇状,多为2个或2个以上长在花序轴的节上(图1A、B)。底部有3~5片由小变大的近卵形苞片包裹,苞片常发生脱落,苞片背面有紫黑色斑点,苞片一般无毛(图1C)。小穗含有两性花3~7朵,平均长3.75 cm,小穗轴节间平均长3.0 mm(图1D)。开花时,往往花枝同一节的小穗开花程度不同,且同一小穗的不同小花之间开花情况和次序也不同。一般由下往上成熟程度递减,即基部小花发育成熟最早,顶部小花发育成熟最晚。小穗轴节间清晰可见,后期会发生逐节断落的现象(图1E)。小穗轴的节间较扁且长度不同,通常基部节间较短,上部节间较长,且同一节间,下部较细,上部较粗(图1F)。

图 1 硬头黄竹小穗的形态解剖Fig. 1 The morphological anatomy of spikelets in Bambusa rigida
2.2 硬头黄竹的小花形态结构
硬头黄竹小花主要由外稃、内稃、浆片、雄蕊和雌蕊五部分组成(图2)。解剖发现,硬头黄竹小花包括内外稃片各1枚,外稃质地较硬,平均长1.25 cm,具多脉,顶端急尖,前端边缘具齿,一般外被纤毛,内侧表面光滑(图2A~D)。内稃对生外稃内侧且短于外稃,有2脊,5脉,上部同样被纤毛(图2E)。浆片3枚,2枚呈半勺形,另外1枚较其它2枚略长,上部边缘有纤毛(图2F)。开花时,浆片吸水膨胀,内外稃张开,此时雄蕊露出小花,因此硬头黄竹又属于开放型花。
硬头黄竹具1枚雌蕊,三分支羽毛状柱头,柱头较长,花柱极短(图2G)。子房呈长卵圆形,较扁,具明显三棱,上部具绒毛,下部光滑(图2H)。雄蕊6枚,未成熟时呈白色,形状整齐,基部呈箭簇状,花药成熟时颜色逐渐变深,且不断长大(图2I)。花药成熟后花丝伸出而垂悬于小花外(图1A)。
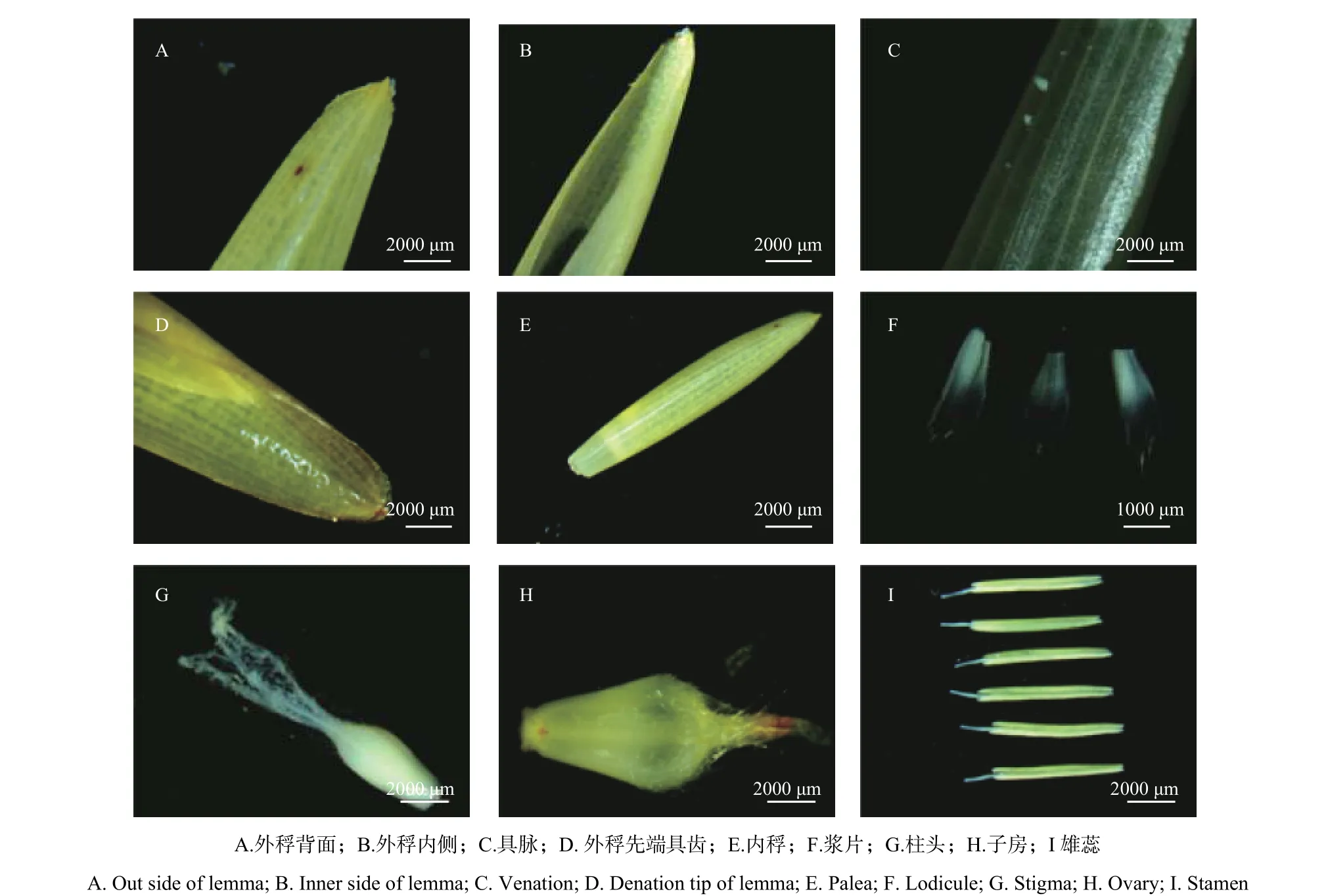
图 2 硬头黄竹小花形态解剖Fig. 2 The morphological anatomy of florets in Bambusa rigida
2.3 硬头黄竹花药的解剖结构与发育
硬头黄竹小穗的每朵小花具6枚花药,每枚花药具有4个药室(图3A)。花药壁的层数随着发育时期不同而有所变化。造孢细胞时期花药壁完全分化,形成四层药壁细胞,从外向内依次是表皮、药室内壁、中层和绒毡层(图3A)。孢原细胞发育为初生造孢细胞后又进行分裂,发育成为次生造孢细胞,此时花药壁为完整的四层结构(图3A)。次生造孢组织游离形成小孢子母细胞(图3B)。在小孢子母细胞时期,绒毡层可能由于营养物质的分泌和消耗,细胞质变淡(图3B)。接着小孢子母细胞开始减数分裂(图3C~E),第一次减数分裂过程中发育为二分体,两个核在同步进行第二次分裂后,会在四个核之间产生细胞壁并且形成四分体,然后形成单核小孢子(图3F)。此时,在药室内仍见到处于减数分解期的小孢子母细胞,这表明硬头黄竹同一花药室内的不同小孢子发育是不同步的(图L)。随着小孢子的体积逐渐增大和细胞质逐渐液泡化,形成许多小液泡,然后小液泡之间互相融合为大液泡,细胞核被大液泡挤到细胞的一端,即单核靠边期(图3G),此时可以在花粉壁上观察到呈凸透镜状的花粉萌发孔,绒毡层发生显著退化,几近消失,只剩下一薄层贴着药室内壁,药室内壁也尚未纤维化形成纤维层。后来液泡逐渐消失,细胞质逐渐充满整个细胞,细胞核进行一次有丝分裂,形成二细胞花粉粒,药室内壁也发生显著纤维化,形成纤维层(图3H)。花粉粒随后进一步发生沉积,形成成熟花粉粒,从而导致花粉粒内的细胞核难以观察到(图3I)。在花粉粒发育成熟后,药室壁连接花丝处比较薄,且未发生纤维化,由于纤维层的作用,进而导致此处药壁纵向裂开,以方便花粉粒释放。在此阶段,花药壁仅剩下表皮细胞和纤维化的药室内壁共两层细胞(图3I)。
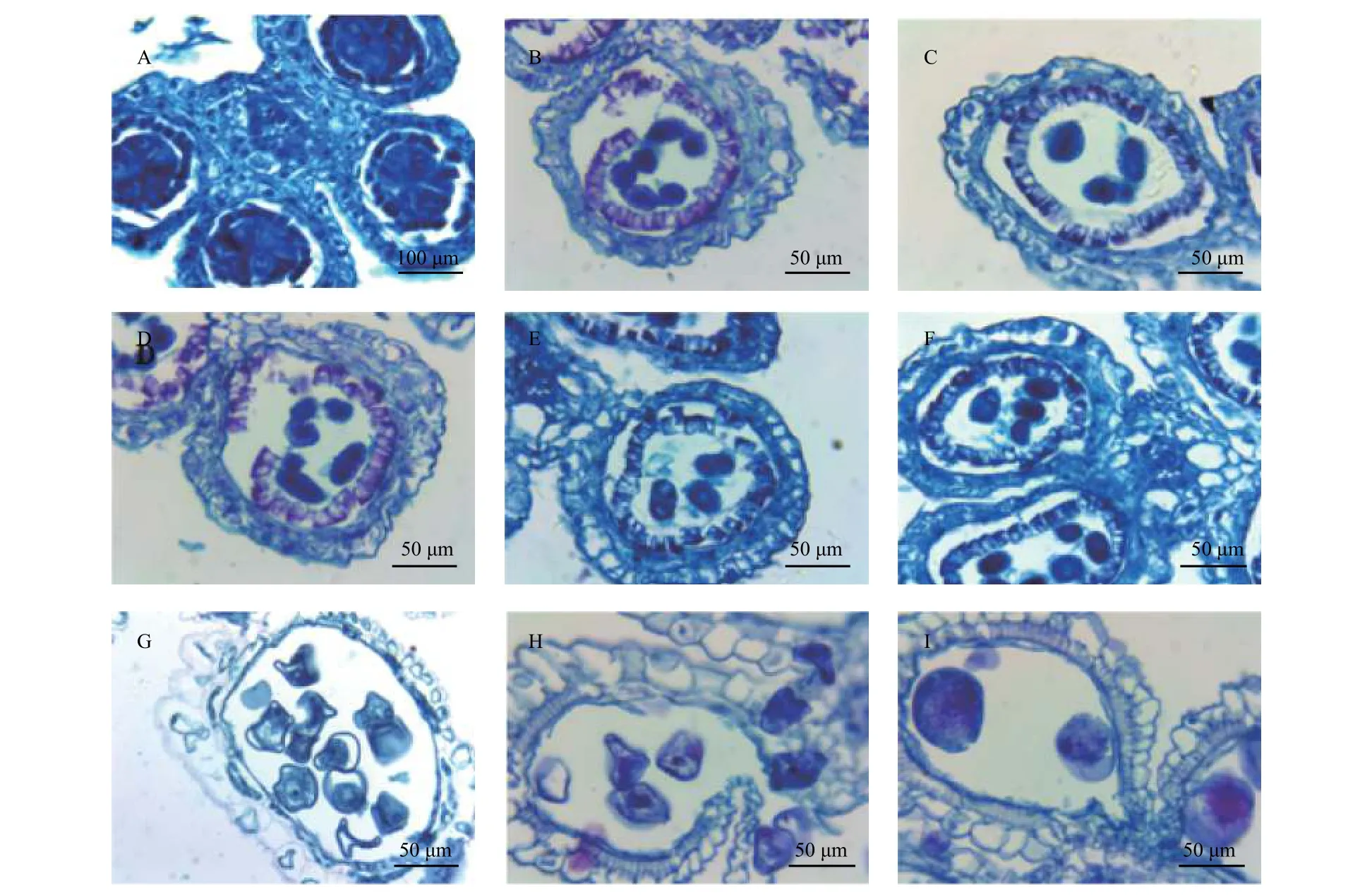
图 3 硬头黄竹花药解剖结构Fig. 3 The anther anatomical structure of Bambusa rigida
2.4 硬头黄竹花药败育
硬头黄竹虽然开花,但结实率极低,几无种子可以收获。通过对花药进行切片观察,可以发现花药出现大量败育的现象。对这些败育的花药进行解剖观察分析,可归纳总结为以下几种类型(图4)。
第一种类型为花药壁发育正常,能形成正常的纤维层,绒毡层亦能退化,药室内仅个别花粉粒发育正常,但其他花粉粒发生收缩等异常现象(图4A)。第二种类型为花药壁发育异常,未能形成纤维层,药室内的小孢子几乎全为空壳花粉粒(图4B)。第三种类型为花药壁也未能形成纤维层,花粉粒全部收缩变形,药室也可能会一起发生收缩(图4C、D),该类型也可能是第二种类型的不同发育阶段。第四种类型为花药壁能形成绒毡层,且绒毡层发生退化,花药壁也未发生纤维化,甚至药室为空药室,未产生任何小孢子(图4E)。第五种类型为花药壁未能够正常分化,花药室也未能产生正常的小孢子(图4F)。
2.5 硬头黄竹子房的解剖结构
硬头黄竹极少开花,也未能收获到发育成熟的果实。在采集硬头黄竹小穗的时候,由于只有一丛竹子中的几株开花,因此能够采集到可用于实验的小穗不多,加上子房极小,很难切到不同发育时期的雌配子体,因此可以观察并用以进行描述的代表性切片较少(图5)。硬头黄竹子房位于雌蕊下端,子房上部外披较硬的纤毛、下部光滑,且有棱边。纵切发现该竹种为侧膜胎座,倒生胚珠(图4A),2层珠被(图4B)。对膨大的子房进行纵切,可偶尔发现发育中的种胚(图4C)。但由于野外未能收获发育成熟的果实,也未在竹丛下部的土壤中发现实生苗,因此部分受精成功的子房未能发育成正常种子的原因并不清楚。而与此形成鲜明对比的是,在校园开花慈竹(Neosinocalamus affinis)竹丛林下的土壤中可以观察到大量存活的实生苗。
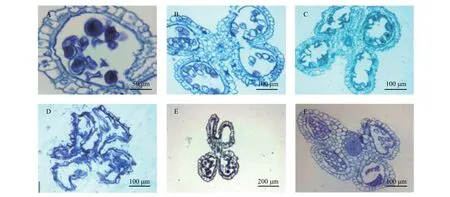
图 4 硬头黄竹各种发育异常的花药Fig. 4 Various kinds of abortive anthers in Bambusa rigida
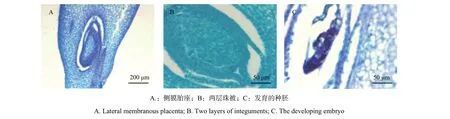
图 5 硬头黄竹子房Fig. 5 The ovary of Bambusa rigida
另外观察发现,开花的硬头黄竹竹丛中少数竹秆开花,并没有全部竹秆一起持续开花,开花后相应的竹秆、竹丛也未死去,仍正常生长,也未再次开花。由于未见到其它地理种源的硬头黄竹开花特征是否与此一致,因此未能明确此特征是否为该竹种的共同特征。但根据杜凡等[25]的总结,将梨藤竹属(Melocalamus)、空竹属(Cephalostachyum)、滇竹属(Oxytenanthera)、思劳竹属(Schizostachyum)、薄竹属(Leptocanna)、牡竹属(Dendrocalamus)、慈竹属(Neosinocalamus)、悬竹属(Ampelocalamus)等归为开花后死亡类型,新小竹属(Neomicrocalamus Keng f.)、箣竹属(Bambusa)、大节竹属(Indosasa)等为开花不死亡类型,箭竹属(Fargesia)和刚竹属(Phyllostachys)则开花死亡和开花不死亡两种情况都有。本研究中硬头黄竹属于箣竹属下的竹种为开花不死亡类型符合该结论。
3 讨论
目前在竹类植物研究中,有关竹类植物的花序一般是遵循McClure[26]和耿伯介先生[27]的定义。McClure[26通过研究中国的竹种Schizostachyum的花序,在1934年首次提出“假小穗”的概念,他对竹类植物的花序进行了分类,认为竹子的花序可以分为:determinate inflorescence(有限制花序)和indeterminate inflorescence(无限制花序)两大类,其区分标准为小穗的基部有无潜伏芽的存在。通常indeterminate inflorescence 的基部苞片内有休眠芽,可以不断地产生新的小穗状,此种类型为假小穗;而determinate inflorescence的基部没有休眠芽,它的各小花是一次性进行开放的。林树燕等[28]通过对翠竹(Pleioblastus fortunei (Van Houtte ex Munro) Nakai)、黄条金刚竹(Sasaella kongosanensis‘Aureostria-tus’)以及‘霞早’绿竹 (Bambusa oldhamii‘Xia Zao’)的花序建成过程进行了观察,发现同一个花序中顶部的小穗首先发育,然后向基部依次发育,由不同位置小穗共同组成的花序为有限花序;而同一小穗上不同部位小花的发育顺序为基部到顶部的顺序依次开放,为无限制花序。因此,林树燕等[28]认为竹类植物的花序应为混合花序。根据McClure[26]和耿伯介[27]的观点,黄条金刚竹和翠竹的花序类型属于有限花序即真花序,根据其定义小穗基部应没有休眠芽。而林树燕等[27]发现,黄条金刚竹顶部小穗发育完成后,其基部又新分化出小穗。因此林树燕等[28]认为竹类植物小穗基部具潜伏芽,应均为有限制花序(假小穗)。本研究中硬头黄竹小穗采集时间较早,采集时并未关注硬头黄竹的花序,因此不予讨论小穗所组成的花序方式。但由于硬头黄竹与林树燕等所报道的绿竹均属同属,因此两者花序应该相同。在硬头黄竹小穗内,小花由基部向顶部逐渐开放的顺序,属于无限制花序,与林树燕等人的观点一致。由于硬头黄竹小穗基部具有潜伏芽,属于传统观点的“假小穗”。
有关竹类植物花粉败育、结实率低的报道并不是特别多,王雨珺等[4]在绵竹花药中发现了大量败育的现象,并对花药败育进行分析,认为花药缺乏正常的造孢细胞、绒毡层中层发育异常、药室收缩变形、花粉粒中空、花粉粒皱缩等多种变异类型导致花药败育。林树燕[29]认为花粉败育及萌发率低、胚囊发育异常、柱头可授性低及自然授粉率低等情况均引起竹类植物的结实率显著降低。庞延军等[30]认为爬竹(Drepanostachyum scandeus (Hsuch et Yi) Keng f. ex Yi)雄蕊发育异常的现象最早出现在雄蕊原基发育时期,发育异常的雄蕊不能形成花粉囊或花粉囊内细胞高度液泡化,无法形成正常的小孢子母细胞。王曙光等[10]认为巨龙竹的小穗中存在营养竞争,致使顶端的小花先发育结实,并先一步成为营养中心,而开花濒死的巨龙竹本身营养物质有限,进而限制其它小花的发育。除此之外,在青丝黄竹[5]、鹅毛竹[9]中均报道过多种败育现象,这些败育现象中与本研究中发现的硬头黄竹药室内的小孢子几乎全为空壳花粉粒、花药壁未能形成纤维层,花粉粒及药室收缩变形及花药壁的绒毡层退化,花药壁未发生纤维化,甚至药室为空药室,未产生任何小孢子等观察到的败育现象相似。因此,花粉败育可能是竹类植物败育过程中发生共同现象,是竹类植物结实率低的共同原因。
硬头黄竹的雌雄异熟的特殊条件导致硬头黄竹结实率降低。雌蕊和雄蕊几乎都不是同时成熟的,导致硬头黄竹不能自花授粉,多是异花授粉现象。而异花授粉现象在一定程度上加大了授粉成功的难度,由此降低了硬头黄竹的结实率。同时,硬头黄竹的开花季节在雨季,雨季花粉粒沾水后导致其传粉性能变差,由于其授粉的方式为风媒,因此在一定程度上也降低了硬头黄竹的授粉几率。在有限的范围内,开花的竹丛少,可授粉的雌蕊又不足的情况下,必然会导致硬头黄竹的结实率下降。
4 结论
硬头黄竹小穗基本结构发育正常,小穗基部具潜伏芽,为假小穗。小穗开花顺序由基部向顶部开放,属于无限制花序。小穗发育正常,包含3~7朵小花,小花为开放型,具内外稃各1片,浆片3枚,雌蕊1枚,雄蕊6枚。雌蕊为三分枝羽毛状柱头,花柱极短。子房长卵圆形,具三棱。子房单室,侧膜胎座、倒生胚珠。花粉粒为二细胞花粉,花药发育后期出现大量败育现象,可总结为5种败育类型。花药败育是导致硬头黄竹结实率低的主要原因。
