低平均光强下波动光对铜绿微囊藻生长的影响
2020-04-06汤加刚李鹏飞何飞飞贺健程
孙 昕 汤加刚 李鹏飞 孙 杰 何飞飞 由 黎 贺健程
(西安建筑科技大学陕西省环境工程重点实验室, 西北水资源与环境生态教育部重点实验室, 西安 710055)
光照是浮游植物生长的重要能量来源, 具有复杂的空间和时间变化模式。水体中浮游植物接受光照的变化分为两种, 一种是水面入射光变化, 这主要由一天中太阳光的光强、气候的变化等引起;另一种是藻类在水中由于湍流、内波等垂直混合作用引起浮游植物在水中位置的改变, 进而影响其接受的光强大小。
目前研究主要集中于两种形式的波动光: 一种是矩形波动光[1,2], 主要由两种不同强度的恒定光不断循环交替产生。另一种是非矩形波动光[3,4],由正弦波动或余弦波动演变而来。
波动光通过影响光合作用和呼吸作用等重要生理过程而影响浮游植物的生长[5,6]。Litchman[7]研究表明在波动光下浮游植物的生长速率变化具有物种特异性, 在低平均光强[25 μmol photons/(m2·s)]条件下, 矩形波动光[15/35 μmol photons/(m2·s)]促进了硅藻Nitzschiasp生长, 抑制了蓝藻Phormidium luridumd和绿藻Sphaerocystis schroeteri的生长, 所以当其作用于浮游植物群落时, 群落组成和结构就会有较大的变化; Litchman[8]还发现在高平均光强[100 μmol photons/(m2·s)]条件下, 矩形波动光[65/135 μmol photons/(m2·s)]组的浮游植物的种类比恒定光照组多; 同时波动光对浮游植物的色素[4]、油脂[9]等合成也有显著的影响, 所以探究波动光对浮游植物的生长影响具有重要意义。
关于不同频率的矩形波动光对浮游植物的研究很多。Liao等[10]发现随着光暗转换频率的增加,其生物量和生长速率也随之增加, Iluz等[11]用自制的生物反应器得出相似的结论。浮游植物在垂直运动过程中, 接受的波动光强可以从完全黑暗到超过1500 μmol photons/(m2·s)的强度, 其经历的光照是由光限制到光抑制的一种渐变过程, 而非瞬变过程, 所以用矩形波动光代替浮游植物垂直运动过程中的光变化有很大的局限性。目前关于非矩形波动光的变化频率(即波动周期)对藻类影响的研究甚少。Nicklishch[12]发现在平均光照相同时, 周期为60min非矩形波动光[1—1000 μmol photons/(m2·s)]下浮游植物的增长速率普遍比周期为30min波动光大, 但尚未针对波动周期对浮游植物影响进行系统性的研究。
浮游植物在自然水体中接受的光照受到表面光照与位置变化的共同作用。在水体表面光强恒定的条件下, 本研究注重浮游植物因内波运动作垂直运动而接受的波动光。通过采用一种新型的基于单片机系统的装置, 分别模拟了内波作用下的波动光强和波动过程的平均光强, 以及对叶绿素荧光技术的应用, 旨在探究波动光的周期性变化对铜绿微囊藻Microcystis aeruginos生长的影响以及其在不同光照条件的光合响应机制。
1 材料与方法
1.1 实验装置
如图 1所示, 实验装置是由外部水浴加热系统、内部培养系统及单片机控制系统三部分组成。外部加热系统由水浴箱、碳纤维潜水加热线和温控系统构成。内部培养系统由培养箱和LED灯组成, 通过恒温水浴箱调控培养箱中藻液温度, LED灯为藻类生长提供光源。单片机系统由单片机、电机驱动片机模块、直流电源、调光驱动器等构成。单片机是整个系统的核心, 通过编写不同时刻的PWM波的占比(范围为0—100%), 可以实现LED光强的周期性变化。
1.2 模拟方法
实验所采用的光是基于水体中出现的内波现象而引起的波动光。在内波作用下藻类位置会发生垂直往复变化; 当水面光强一定时, 水下光照强度随水深增加而不断衰减, 所以藻类接受的光强也会随之变化。在内波条件下, 藻类的运动可以理想化为如式(1)所示的简单的波动方程:

式中,Zi为内波产生的水深,A为波动振幅,T为波动周期, 本实验中Zi取7.9 m,A取2 m,T分别取10min、1h和6h。
根据Lambert-Beer光衰减特性方程, 在任意水深Z处的光强可用式(2)表示如下:

式中,Iz为水深为Z处的光强,i0为水体表面光强取1000 μmol photons/(m2·s),k为光的衰减系数取0.5/m。
将式(1)与式(2)合并, 得水深Zi处藻类在内波作用下接收的波动光FL形式, 如式(3)
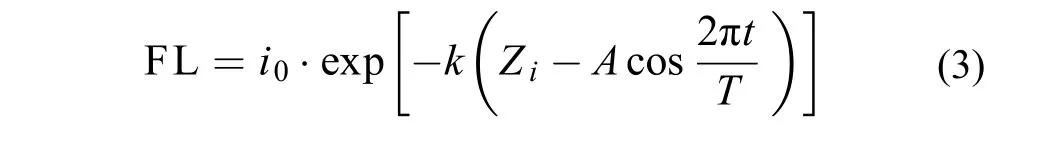
图 2表示在波动周期为10min的波动光强条件下, 一个周期内光照强度随时间的变化关系。在0时, 藻类受内波作用达到波动的最高点, 即水深5.9 m处, 光强为52.34 μmol photons/(m2·s); 在300s时, 藻类达到波动最低点, 即水深9.9 m处, 光强为7.08 μmol photons/(m2·s)。
在一个波动周期内, 水深Zi处藻类在垂向波动过程中接受的平均光强AL可用式(4)计算如下:


图 1 基于单片机的光强控制实验装置Fig. 1 The experimental device based on a single-chip system for controlling light intensity
由式(4)可知, 波动光在一个周期内的平均光强AL大小与其周期无关。根据上述条件, 可计算出本实验中的平均光强AL≈25 μmol photons/(m2·s)。

图 2 理论光强变化机制Fig. 2 Theoretical light intensity change mechanism
通过光照强度测量和单片机编程的调节, 可以实现不同特性的光照条件。理论上波动光时刻都在变化, 但是由于单片机程序的局限性无法实现这种变化频率, 本实验模拟的实际光强的变化频率为0.1 Hz(图 3)。

图 3 实际光强调节机制Fig. 3 Actual light intensity adjustment mechanism
1.3 实验条件
实验选用的铜绿微囊藻(Microcystis aeruginosaFACHB-569)购买于中国科学院淡水藻种库。参考文献[13], 铜绿微囊藻初始藻密度为2.0×106cells/mL, 培养基为BG11, 藻液温度为25℃。在实验装置中单格反应器中所盛藻液体积为1 L, 藻液厚度为4 cm左右, 可以减少藻类自身互遮影响。
将铜绿微囊藻置于恒定光照和波动光照等四组不同光照条件下培养, 并且每组光照均设置3个平行样。三组不同周期波动光照, 分别为10min FL、1h FL和6h FL, 其中10min FL表示波动周期为10min的波动光强, 其余类同。三种波动光照条件下的平均光强相同, 均为25 μmol photons/(m2·s)和一组恒定光强AL组[光强为25 μmol photons/(m2·s)]。在所有光照条件的实验中, 均采用12h∶12h的光暗(L∶D)培养。
1.4 藻密度
每隔一天取适量藻液, 采用血球计数板计数法测定铜绿微囊藻密度。
1.5 比增长速率
铜绿微囊藻的比增长速率计算公式见(5),

式中C1为铜绿微囊藻培养t1天的藻密度,C0为铜绿微囊藻培养t0天的藻密度
1.6 类胡萝卜素
采用文献[14]所述方法测定类胡萝卜素含量。在紫外可见分光光度计下测定待测样在波长分别为663、645和450 nm的吸光值, 利用式(6):

待测样的制备: 取藻液5 mL放入离心管中, 在离心机上以9000 r/min离心15min, 离心后弃掉上清液, 向离心浓缩沉淀中加入80%丙酮溶液, 并均匀混合。然后将离心管用锡箔纸裹住, 放入4℃冰箱中静置24h, 进行脂溶性色素提取,色素提取后, 再将其在8000 r/min离心10min, 上清液即为待测样。
1.7 荧光参数
取1.5 mL藻样于离心管中, 暗适应10min后使用调制叶绿荧光成像测试系统(Imaging-pam, 德国WALZ)测定。
1.8 数据的处理和分析
采用Excel 2003和SPSS 16.0软件对数据进行统计分析, 使用Origin 8.5数据图绘制。
2 结果
2.1 藻密度和比增长速率
从图 4中可以看出, 铜绿微囊藻在不同光照条件下的生长趋势基本一致。四组光照条件下的铜绿微囊藻藻密度在培养末期差异显著(P<0.05), 其中平均光强AL组较波动光FL组增殖较慢且提前进入稳定期。在22d时, 6h FL、1h FL和10min FL组的铜绿微囊藻藻密度相对于AL组分别增加了28.3(P<0.05)、18.2%(P<0.05)和7.7%(P>0.05)。
在低平均光强下, 在不同光照条件下铜绿微囊藻的比增长速率差异显著(P<0.05), 6h FL、1h FL和10min FL组的铜绿微囊藻比增长速率相对于AL组分别增加了10.1(P<0.05)、6.8%(P<0.05)和3.0%(P<0.05, 图 5)。
2.2 单个细胞类胡萝卜素含量
在低平均光强下, 单个铜绿微囊藻细胞所含的类胡萝卜素含量随着波动光周期的增大而显著降低(P<0.05), 在第10天时, 6h FL、1h FL和10min FL组的单个细胞类胡萝卜素含量比AL组分别降低了18.1%(P<0.05)、16.7%(P<0.05)和6.9%(P<0.05,图 6)。
2.3 叶绿素荧光参数(Fv/Fm、ETR、NPQ)
从图 7可以看出, 在四种光照条件下, 铜绿微囊藻的最大量子产量Fv/Fm值在前4d都是呈现逐渐增大趋势, 这是由于初期藻细胞浓度较低, 细胞间的互遮效果不明显, 保证了单位藻体的最佳光强需求,从而造成了在藻类生长前期Fv/Fm值的增大。在低平均光强下, 铜绿微囊藻Fv/Fm值随着波动光周期的增大而显著升高(P<0.05), 其与藻密度和比增长速率相对大小关系一致。
从图 8中可以看出, 每组光照条件的相对电子传递速率(rETR)随光合有效辐射(PAR)强度的增大而增大。当光照强度大于110 μmol photons/(m2·s)时, 不同光照条件下的rETR差异显著(P<0.05), 且6h FL组拥有最大的rETR值, 其次为1h FL、10min FL、AL组, 与Fv/Fm值相对大小关系一致。
从图 9可以发现, AL组的NPQ一直处于缓慢上升的状态, 其6h的NPQ值相对于0时增加了104%(P<0.05)。6h FL组的NPQ值在0、3h和6h时值分别为0.025、0和0.023, 其趋势在一个周期内先减小后增大, 这表明铜绿微囊藻在三种机制下的热耗散能力有差异。各组光机制下热耗散NPQ平均值大小关系为: NPQ(AL)>NPQ(1h FL)>NPQ(6h FL)。

图 4 不同光照条件铜绿微囊藻的生长曲线Fig. 4 Growth curves of Microcystis aeruginosa under different light conditions

图 5 不同光照条件对铜绿微囊藻比增长速率的影响Fig. 5 Effects of different light conditions on the specific growth rate of Microcystis aeruginosa

图 6 不同光照条件对单个细胞类胡萝卜素的影响Fig. 6 Effects of different light conditions on carotenoids in single cells
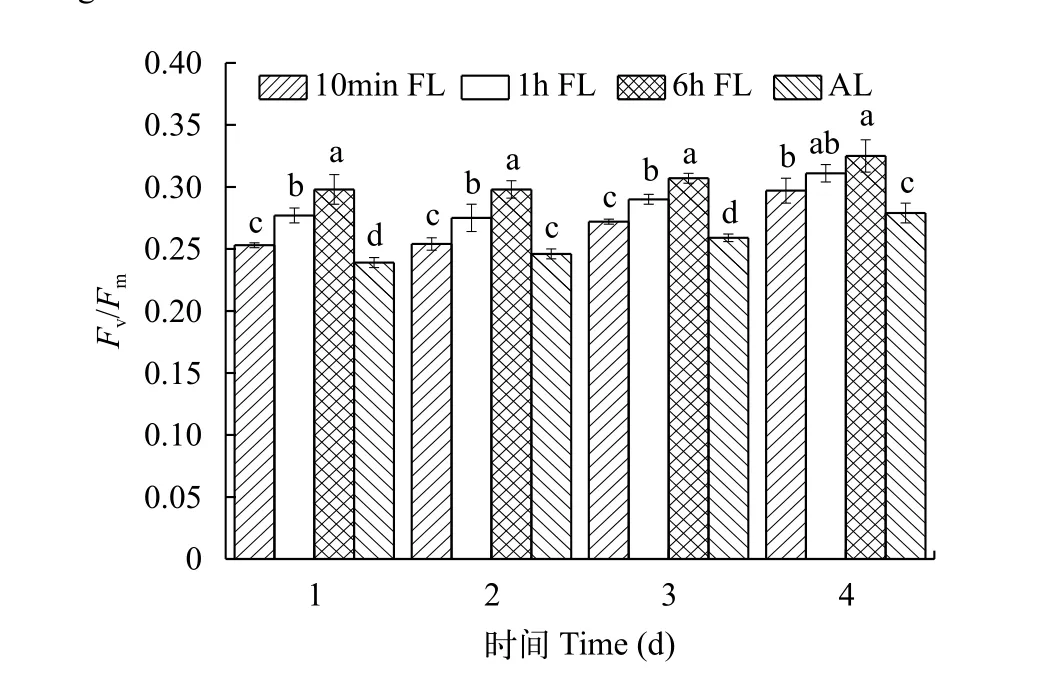
图 7 不同光照条件下Fv/Fm值变化Fig. 7 Changes in Fv/Fm values under different light conditions
3 讨论
类胡萝卜素广泛存在于藻类和植物中, 其重要的功能就是通过猝灭三线态及单线态氧的能量, 保护光合系统正常工作。有关研究表明, 藻类在不利的条件下为了保护组织不受伤害而提高类胡萝卜素的合成[15—17]。本研究结果显示各组波动光下的单位类胡萝卜含量差异显著(P<0.05), 且随着波动光周期的减小而增大。藻类的中间电子受体在低光或者无光的环境下可再生和再氧化, 从而改善电子传递过程, 而连续的恒定光照会损伤质体醌中的中间电子受体[10,18]。为了维持细胞自身的生长, 铜绿微囊藻通过增加类胡萝卜素的合成量以缓解由于叶绿素分子受到光损伤而导致的光合机能的衰减[16]。由于波动光的特殊性, 其在高光和低光状态的不断转换, 减少对藻类的持续性光损伤。波动光周期越小, 高光到低光变化频率越大, 这种快速的环境改变也会对藻类产生胁迫, 相应地, 也会提升类胡萝卜素含量的合成来保护自身的光组织。

图 8 不同光照条件下相对电子传递速率随光强变化Fig. 8 Changes in the relative electron transfer rate with light intensity under different light conditions

图 9 非光化学猝灭的时变化Fig. 9 Temporal variation in non photochemical quenching
叶绿素荧光技术是近年来研究植物光合作用机理的一种重要手段。如图 4、图 7、图 8, 虽然在前4天铜绿微囊藻密度差异不显著(P>0.05), 但Fv/Fm值和rETR值差异明显, 因为当外界条件变化时, 会在很短的时间反应在藻类的生理效应上。在本实验中波动光强组的Fv/Fm值和rETR值都显著大于平均光强组, 这与Wedchaparn等[19,20]研究结果一致。微藻吸收的光能, 一部分用于光合作用, 另一部分多余的能量以热能的形式耗散掉。光照强度会影响藻的非光化学淬灭(NPQ), 而且随着光照强度升高, NPQ会逐渐增大[21—23], 本实验也得出相似结论。各组光照条件下NPQ大小关系与Fv/Fm、rETR值相反。这表明周期越小的波动光卡尔文循环的活性受抑制增强, 铜绿微囊藻为了保护自身,潜在的热耗散能量增加。
孙昕等[24]研究发现在0—20000 lx的波动光抑制了铜绿微囊藻的生长, 因为藻类在高平均光强波动时, 其接受的绝大部分光照已经超过最适光强,既而其生长受到抑制。而本实验是在低平均光强条件下进行, 研究结果表明相比于恒定光照, 铜绿微囊藻在波动光下能够更好地调节自身光合作用机制去利用光能。产生这样的原因可能来自两方面: 一方面是恒定光照会产生活性氧中间体, 这可能会产生光抑制现象, 从而限制了微藻生长[25]; 另一方面由于连续的恒定光照损伤了质体醌中的中间电子受体, 限制了光合作用电子传递的关键步骤[17]。由于波动光是从高光到低光或者无光状态不断的转换, 避免了恒定光照状态, 增强了光合作用的电子传递。同时波动光周期越小对电子传递限制越明显, 原因可能是高光到低光变化频率越大, 导致藻细胞频繁适应高光与低光之间转换的机制, 间接地损害了光反应系统, 从而降低了光合作用效率。
