绵羊RXRG基因的生物信息学分析
2020-04-01张司龙张小雪宋其志王维民
张司龙 张小雪 宋其志 王维民


摘要:以绵羊RXRG基因为目的基因,利用生物信息学软件预测其结构和功能。结果表明,绵羊RXRG基因编码463个氨基酸,开放阅读框长度为1 392 bp,起始密码子位于228 bp处,终止密码子位于1 619 bp处。RXRG基因编码蛋白的相对分子质量为50 845.19 Da,等电点为7.55,在氨基酸组成中亮氨酸所占比率最高,色氨酸占比最低。亚细胞定位主要位于细胞核中,不属于分泌蛋白;不存在信号肽序列;存在两个保守结构域,并且为疏水性蛋白,二级结构主要以α螺旋和无规则卷曲为主,三级结构主要由无规卷曲缠绕折叠形成。
关键词:绵羊;RXRG基因;生物信息学分析
中图分类号:S826 文献标志码:A 文章编号:1001-1463(2020)02-0031-07
Abstract:In this study, with the sheep RXRG gene as the target gene, the structure and function of sheep RXRG gene were predicted by bioinformatics software. The RXRG gene of sheep encodes 463 amino acids, the length of open reading frame is 1 392 bp, the initial codon is 228 bp, and the termination codon is 1 619 bp. The relative molecular weight and isoelectric point of RXRG protein were 50 845.19 Da and 7.55 respectively. Leucine accounted for the highest proportion of amino acid composition and tryptophan accounted for the lowest proportion. Subcellular localization is mainly located in the nucleus, not in the secretory protein;there is no signal peptide sequence;there are two conservative domains, and they are hydrophobic proteins. The secondary structure is mainly αshelix and irregular curl, and the third structure is mainly formed by random curl winding and folding.
Key words:Sheep;RXRG gene;Bioinformatics analysis
視黄酸是一种脂溶性的小分子物质,在细胞分化、上皮细胞生长、视觉和组织维持、胎儿发育和繁殖等过程中发挥着重要作用[1 ]。视黄酸受体(Retinoic acid receptor,RAR)和视黄素X受体(Retinoid X receptor,RXR)是视黄酸受体家族的主要成员,RAR、RXR由3种不同的基因RXRA、RXRB和RXRG编码,形成了RXRA、RXRB,RXRG等多种类型的受体[2 - 3 ]。RXRG基因作为配体激活的转录因子,结合到靶基因的特定应答序列上,调节基因的转录表达[2 ],是细胞分化和组织形态发生的主要调节因子。RXRG基因的研究多在黑猩猩、猕猴、狗、奶牛、大鼠、鸡、斑马鱼、青蛙等方面,而关于绵羊RXRG基因的研究较少。国内外研究表明,RXRG基因不仅在动物妊娠的关键时期表达,还对具有高产性能猪种的窝产仔数具有明显的加性效应,对于高繁殖力的绵羊品种具有显著影响[4 ]。因此,对RXRG基因进行深入研究,利用生物信息学相关软件和工具,对不同物种RXRG基因系统发育和绵羊RXRG基因CDS区序列蛋白质的理化性质、二级结构及多参数预测、蛋白质跨膜结构、信号肽预测、亚细胞定位和三级结构等进行分析,旨在寻找与双胎或者多胎性状有关的遗传标记位点,为培育高繁殖力绵羊品系提供理论依据,为绵羊RXRG基因结构与功能研究提供参考[5 ]。
1 材料与方法
1.1 序列来源
数据资料来源于NCBI网站的GenBank数据库,包括绵羊(XM_012185408.2)、瘤牛(XM_019986703.1)、黑猩猩(XM_513962.5)、人(NM_006917.5)、家鼠(NM_009107.3)、猪(NM_001130213.1)、马(XM_023640805.1)和鸡(NM_205294.1)8个物种的mRNA序列(括号内为Gen Bank登录号)。
1.2 方法
绵羊RXRG基因开放阅读框采用NCBI的开放阅读框查找器程序分析;采用DNA生物编辑器(Bioedit及DNA Star)分析软件预测RXRG基因编码产物的理化性质;多序列比对及同源性分析采用分子生物学综合应用软件(DNAMAN);采用蛋白质亚细胞定位预测工具(PSORT)进行亚细胞定位。在蛋白质的分析预测方面采用信号肽预测工具(Signalp3.0软件)进行蛋白潜在信号肽剪切位点的预测,采用捆绑混合隐马尔可夫模型[tied-mixture hidden Markov models(TMHMM)]程序进行跨膜螺旋区域的预测,采用简单模块化架构研究工具(SMART)软件进行蛋白保守结构域分析;二级结构采用蛋白质二级结构预测服务器(Jpred)分析预测;采用瑞士模型(Swiss-model)软件分析蛋白三级结构;在蛋白亲疏水性分析方面采用疏水性分析(ProtScale)程序[4 - 6 ]。
2 结果与分析
2.1 绵羊RXRG基因开放阅读框
开放阅读框的分析结果如图1所示,绵羊RXRG基因推测编码463个氨基酸残基,其基因序列长度为1 392 bp,起始密码子位于228 bp处,终止密码子位于1 619 bp处。
2.2 绵羊RXRG编码产物的理化性质
蛋白质的理化性质分析包括对其相对分子质量、氨基酸组成以及等电点的理化性质分析[7 ]。绵羊RXRG基因编码产物的理化性质结果(图2)表明,该基因共编码463个氨基酸残基,相对分子质量为50 845.19 Da,其氨基酸残基中亮氨酸所占比重最高,为9.9%;其次为丝氨酸,所占比例为9.5%,甘氨酸所占比例为7.3%;色氨酸占比最低,仅为0.4%。
2.3 绵羊RXRG基因蛋白亚细胞定位
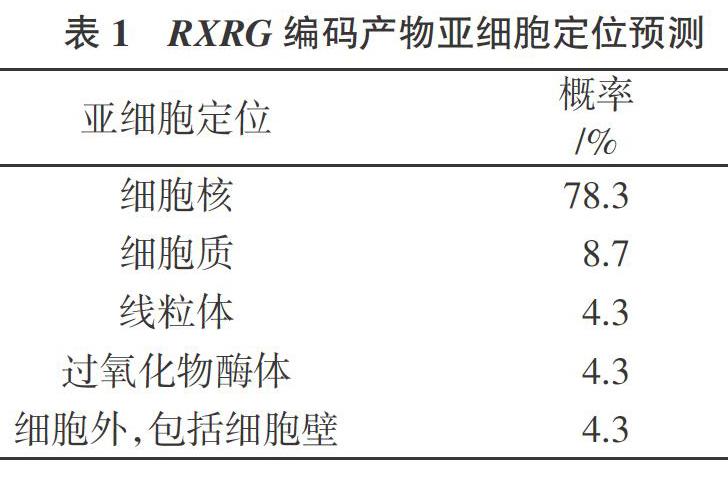
从蛋白质亚细胞定位预测工具程序预测得到的绵羊RXRG基因蛋白亚细胞定位结果(表1)可以看出,绵羊RXRG蛋白的亚细胞大多数分布在细胞核,其可能性为78.3%。而在其他位置分布的可能性所占概率较小。其中,分布于细胞质的可能性为8.7%,分布于线粒体的可能性为4.3%,还有4.3%分布在过氧化物酶体。由此推断,绵羊RXRG基因主要在细胞核中发挥生物学作用,其次在细胞质中发挥作用。
2.4 绵羊RXRG编码产物序列同源性
通过NCBI数据库找到绵羊、瘤牛、猪、马、黑猩猩、人、家鼠、鸡八种动物DNA序列,并对其进行同源性分析(图3),可以看出,绵羊与他们之间的亲源性分别为98%、93%、92%、91%、91%、86%、73%。由系统发育树可看出在这8种动物中绵羊RXRG基因与瘤牛亲缘关系较近,与鸡的亲缘关系最远,RXRG基因在哺乳动物间的保守性较高(图4)。
2.5 绵羊RXRG蛋白潜在信号肽剪切位点预测
通过分析预测绵羊RXRG蛋白潜在信号肽剪切位点的情况可以得知基因编码的产物是否是分泌蛋白和跨膜蛋白,以及跨膜蛋白的基本信息。从图5可以看出,信号肽序列是存在于分泌蛋白基因编码序列中起始密码子之后的一段富含疏水氨基酸多肽的序列,绵羊RXRG蛋白氨基酸在23的位置出现最大C值为0.119,截止点为0.32;在1的位置出现最大S值为0.824,截止点为0.87;在1-19的位置平均出现S值为0.560,截止点为0.48;在20的位置出现最大Y值为0.178,截止点为0.33;在1-19的位置出现D值为0.369,截止点为0.43因此推断绵羊RXRG基因的编码产物属于分泌性蛋白。位置19和20之间最可能的裂解位为CGG-CT。
2.6 绵羊RXRG蛋白跨膜区域结构预测
从图6可以看出,通过使用TMHMM软件对绵羊RXRG蛋白跨膜区域结构进行预测分析,发现该基因所编码的蛋白没有跨膜区域结构。
2.7 绵羊RXRG蛋白保守结构域分析
由Smart软件分析结果得出,绵羊RXRG基因在第136~207位、第271~430位氨基酸残基之间存在跨膜区,在76~89的位置上出现了低复杂性区域(图7)。
2.8 綿羊RXRG蛋白亲疏水性分析
从图8看出,蛋白质亲疏水性研究发现该基因编码蛋白疏水性达到最大值的位点有很多,且最大值为4.000,其中Arg、Leu、Ser的单个值最高,达到了6.000。此外,亲疏水性分析图中最小值为1.667,分析其基因得到的结果全部为正值,可见其并无亲水性,从而得出该基因编码的蛋白属于疏水性蛋白。
2.9 绵羊RXRG蛋白二级结构的预测
通过Jpred软件进行蛋白质二级结构的预测分析(图9)可知,绵羊RXRG蛋白二级结构中有大部分结构为无规卷曲,其次为α螺旋,β折叠数量较少。其结果为10个α螺旋,4个β折叠,其余全是无规卷曲结构。
2.10 绵羊RXRG蛋白三级结构预测与分析
由图10可知,RXRG基因编码蛋白的三级结构主要由无规卷曲折叠缠绕形成。
3 小结与讨论
绵羊RXRG基因的ORF长度为1 392 bp,编码463个氨基酸残基,起始密码子位于228 bp处,终止密码子位于1 619 bp处。对其理化性质分析发现组成最多的氨基酸是亮氨酸,所占比例为9.9%,其次,丝氨酸占比为9.5%,色氨酸组成所占比例最少,为0.4%。在系统发育树中绵羊RXRG基因同源性与瘤牛关系最近,为98%,其次为猪和马,在所分析的动物中绵羊与鸡的亲缘关系最远。RXRG基因编码蛋白潜在信号肽剪切位点的预测最大C值为0.119,截止点为0.32;最大S值为0.824,截止点为0.87;最大Y值为0.178,截止点为0.33;在1-19的位置平均出现S值为0.560,截止点为0.48;在1-19的位置出现D值为0.369,截止点为0.43;位置19和20之间最可能的裂解位点是CGG-CT。绵羊RXRG基因编码蛋白质为疏水性蛋白,其疏水性最大值为4.000,最小值为1.667,该基因编码的蛋白没有跨膜螺旋结构,α螺旋为主,三级结构主要为无规卷曲,绵羊RXRG蛋白的亚细胞分布在细胞核的可能性最大,为78.3%。
RXRG基因是调节细胞分化和组织形态发生的重要因子,在提高羊生产力方面有重要影响,李华振等[4,8 ]在绵羊繁殖轴相关组织RXRs基因的表达及RXRA基因多态性与季节性发情和绵羊基因多态性与产羔数方面进行了研究,Shaoze Cheng等[9 ]在RXRG调节鸡原始生殖细胞分化方面做过研究。此外,黄萌等[1 - 2,10 ]做过牛RXRG基因遗传变异与双胎性状的关联性分析方面已有研究。在国外还有关于RXRG基因在黑猩猩、斑马鱼以及其他不同功能方面的研究(全外显子鉴定家族分离性催乳素瘤中的RXRG和TH种系变异)[11 ],以及生物信息学的处理理论研究方法等[12 - 13 ],这些研究都表明视黄素受体基因在提高繁殖力方面具有很大潜力。本研究显示,RXRG基因为1 392 bp长度的基因序列,编码463个氨基酸残基,其主要的作用区域是细胞核,其理化性质预测中氨基酸组成中亮氨酸所占比例最大,占比最少的氨基酸是色氨酸。通过系统发育树可以知道绵羊、瘤牛、马、猪在RXRG基因这一方面有比较近的亲缘关系,和鸡的亲缘关系最远,说明RXRG基因在哺乳动物间保守性较高。绵羊RXRG基因编码产物属于分泌蛋白,位置19和20之间最可能的裂解位点是:CGG-CT,该基因编码的蛋白为疏水性蛋白,其保守结构域有两个功能区域,其二级结构多为无规卷曲和α螺旋,其三级结构主要以无规卷曲为主,并且没有发现RXRG基因编码蛋白质产物的跨膜区域结构。
参考文献:
[1] 黄 萌,许尚忠,昝林森,等. 牛RXRG基因遗传变异与双胎性状的关联分析[J]. 遗传,2008(2):190-194.
[2] 黄 萌,许尚忠,昝林森,等. 牛RXRG基因cDNA的克隆及生物信息学分析[J]. 西北农林科技大学学报(自然科学版),2008(11):1-5;10.
[3] ALEXANDRA TALLAFUSS,LAURA A. HALE,YI-LIN YAN,et al. Postlethwait. Characterization of retinoid-X receptor genes rxra,rxrba,rxrbb and rxrg during zebrafish development[J]. Gene Expression Patterns,2005, 6(5):556-565.
[4] 李华振,狄 冉,郭晓飞,等. 绵羊繁殖轴相关组织RXRs基因的表达及RXRA基因多态性与季节性发情之间的关系[J]. 农业生物技学报2019,27(8):1401-1409.
[5] 张小雪,李发弟,王维民. 绵羊STMN2基因生物信息学分析[J]. 甘肃农业科技,2016(7):58-61.
[6] 王 伟,杨巧丽,谢开会,等. 合作猪HMOX1基因克隆及生物信息学分析[J/OL]. 基因组学与应用生物学,2019[2019-04-29]. http://kns.cnki.net/kcms/detail/45.1369.Q.2019 0426.1608.006.html.
[7] 张小雪,李发弟,王维民. 绵羊ANXA10基因生物信息学分析[J]. 甘肃农业科技,2016(6):1-4.
[8] 王 辉. 中国美利奴羊RXRG基因多态性与产羔数的关联性分析[C]//中国畜牧兽医学会畜禽遺传标记分会. 第十二次全国畜禽遗传标记研讨会论文集. 北京:中国畜牧兽医学会,2010.
[9] SHAOZE CHENG,MAN WANG,YINGJIE WANG,et al. RXRG associated in PPAR signal regulated the differentiation of primordial germ cell[J]. Journal of Cellular Biochemistry,2018,119(8):6926-6934.
[10] 刘 宇,何向东,夏 忆,等. 牦牛MSMB基因的克隆及其生物信息学分析[J]. 中国草食动物科学,2019,39(5):1-5;22.
[11] FLAVIA M. MELO,PATR?魱CIA P. COUTO,ALLEN E. BALE,et al. Whole-exome identifies RXRG and TH germline variants in familial isolated prolactinoma[J]. Cancer Genetics,2016,209(6):251-257.
[12]ZHENG GAO,JONATHAN TERHORST,CRISTOPHER V. VAN HOUT,et al. U-PASS:unified power analysis and forensics for qualitative traits in genetic association studies[J]. Bioinformatics,2020,36(3):974-975.
[13] GOH KELVIN G K,MORIEL DANILO G,HANCOCK STEVEN J,et al. Bioinformatic and molecular analysis of inverse autotransporters from escherichia coli[J]. mSphere,2019,4(4):e00572-e00591.
(本文责编:陈 伟)
